Nimetatakse võrkkesta fotoretseptorite elektrilise koguaktiivsuse rekord. Fotokeemilised reaktsioonid võrkkesta retseptorites. Võrkkesta kihtide struktuur ja funktsioonid
Suurus: px
Alusta näitamist lehelt:
ärakiri
1 Jooksvad testid rubriigis ANALÜÜSIJATE FÜSIOLOOGIA (SENSORSÜSTEEMID) 1. Analüsaatorite üldfüsioloogia 1. Mõiste "analüsaator" võeti füsioloogiasse esmakordselt kasutusele 1909. aastal: a) N.E. Vvedenski b) A.A. Ukhtomsky c) I.P. Pavlov d) Ch. Sherrington 2. Analüsaator on ühtne süsteem, mis sisaldab: a) sensoorseid organeid b) perifeerseid retseptoreid, juhtsektsioon ja tsentraalkoore sektsioon c) perifeerseid retseptoreid, konduktoreid ja keskkoore sektsioone, tagasiside reguleerimise süsteemi d ) juhisektsioon ja tsentraalne kortikaalne sektsioon 3. Spetsiaalsed struktuurid, mis tajuvad stiimuli toimet: a) sünapsid b) sensoorsed süsteemid c) retseptorid d) analüsaatorid 4. Analüsaator ei sisalda: a) retseptori aparaati b) radu c) retikulaarset moodustumine d) keskus ajukoores 5. Stiimuli muundumist närviimpulssiks retseptoris nimetatakse: a) esmaseks kodeerimiseks b) sensibiliseerimiseks c) dekodeerimiseks d) kohanemiseks 6. Stiimuli tugevus on kodeeritud ajukoores. neuron: a) impulsi sagedus b) impulsi kestus c) impulsi amplituud 7. Väliskeskkonna mõju elementaarne madalam analüüs toimub: a) Retseptoris b) Retikulaarses formatsioonis c) Juhtimisradades d) Ajukoores 8. Kõrgeimas peen analüüs väliskeskkonna mõju kohta inimestele toimub: ajus d) ajukoores
2 9. Analüsaatorite interaktsiooni kõrgeim tase: a) bulbar b) vars c) kortikaalne d) taalamus 10. Retseptorid, mis on spetsialiseerunud mitut tüüpi stiimulite tajumiseks: a) polümodaalsed b) efektor c) sensoorne d) spetsiifilised 11. Kontakt retseptorite hulka kuuluvad retseptorid: a) haistmis- b) maitsmis- c) kuulmis- d) visuaalsed 12. kauged retseptorid hõlmavad retseptoreid: a) taktiilsed retseptorid b) valu c) maitsetundlikkus d) kuulmis- 13. interoretseptorid on: a) proprioretseptorid b) vistseroretseptorid c) fotoretseptorid d) Vestibuloretseptorid 14. Kontaktretseptorid hõlmavad retseptoreid: a) Puuteretseptorid b) Haistmisretseptorid c) Vestibuloretseptorid d) Fotoretseptorid 15. Kaugemad retseptorid hõlmavad retseptoreid: a) Maitseretseptorid b) Fotoretseptorid c) Puutetundlikud d) Valu 16. Primaarsed sensoorsed retseptorid on: a) maitsepungad b) sisekõrva karvarakud c) taktiilsed retseptorid d) võrkkesta fotoretseptorid
3 17. Sekundaarsete sensoorsete retseptorite hulka kuuluvad: a) intrafusaalsed lihaskiud b) võrkkesta fotoretseptorid c) puutetundlikud d) haistmisvõime 18. Retseptoripotentsiaalil on iseloom: a) leviv b) lokaalne 19. Milline elektriprotsess registreeritakse esmalt primaarsetes sensoorsetes retseptorites. ? a) retseptori potentsiaal b) generaatori potentsiaal c) aktsioonipotentsiaal 20. Sekundaarsete sensoorsete retseptorite poolt kõige sagedamini sekreteeritud neurotransmitter: a) atsetüülkoliin b) histamiin c) serotoniin d) norepinefriin 21. Retseptori selektiivne tundlikkus teatud stiimuli toime suhtes nimetatakse: a) spetsiifilisus b) akommodatsioon c) erutuvus d) kohanemine 22. Retseptorite kohanemisvõimet pidevalt toimiva stiimuliga nimetatakse: a) majutuseks b) modaalsuseks c) adaptatsiooniks d) retseptorite erutuvuse kodeerimiseks c) retseptorite erutatavuse suurenemine 24. Impulsside sagedus retseptorites nende kohanemise protsessis: a) väheneb b) ei muutu c) suureneb 25. Retseptorites puudub kohanemisomadus: a) Taktiilsed retseptorid b) Maitseretseptorid c) Proprioretseptorid d) Haistmisretseptorid
4 26. Retseptorid, millel praktiliselt puudub kohanemine, on: a) temperatuur b) vestibulaarne c) maitse d) taktiilne 27. Inimese välisanalüsaator on analüsaator: a) Motoorne b) Haistmine c) Vestibulaarne d) Interotseptiivne on analüsaator. analüsaator: a) haistmine b) maitse- c) motoorne d) nahk 29. Inimese välisanalüsaator on analüsaator: a) Vestibulaarne b) Motoorne c) Interotseptiivne d) Maitse 30. Inimese väliste analüsaatorite hulka ei kuulu analüsaator. : a) Vestibulaarne b) Kuulmine c) Visuaalne d) Nahaanalüsaator 31. Inimese sisemised analüsaatorid ei sisalda analüsaatorit: a) interotseptiivne b) Vestibulaarne c) kuulmine d) Motoorne 2. Visuaalse analüsaatori füsioloogia 32. Analüsaatori abiaparatuur. silm ei hõlma: a) silmamuna lihaseid b) miimikalihaseid c) pisaraaparaati d) kaitsevahendeid (kulmud, ripsmed, silmalaud) 33. Silma motoorne aparaat sisaldab tahtelihaseid: a) Viis b) Kuus c) Seitse d) Kaheksa
5 34. Silma võrkkestas on umbes: a) 7 miljonit b) 65 miljonit c) 130 miljonit d) 260 miljonit 35. Millised retseptorid moodustavad võrkkesta kollase laiku? a) Vardad b) Käbid 36. Võrkkesta perifeerias on rohkem: a) koonused b) vardad 37. Silma päeva- ja värvinägemise aparaat on: a) Vardad b) Koonused c) Ganglionrakud d) Bipolaarsed rakud 38. Silma hämaras nägemise aparatuuriks on: a) bipolaarsed rakud b) ganglionrakud c) vardad d) koonused 39. Visuaalse analüsaatori retseptoris retseptori potentsiaali tekkimise ajal membraan: a. ) repolariseerib b) depolariseerib c) hüperpolariseerib 40. Nägemisnärvi silmamunast väljumise kohta nimetatakse: a) pime kollatähnik b) fovea c) terminaalne rada d) kollatähni 41. Milliste võrkkesta rakkude aksonid moodustavad nägemisnärvi? a) Amakriinne b) Horisontaalne c) Bipolaarne d) Ganglioniline 42. Retseptorite kogumit, mille ärritus põhjustab ühe võrkkesta ganglionraku ergastamist, nimetatakse: a) retseptiivne väli b) pimeala c) kollane laik d) fovea fovea.
6 43. Visuaalse analüsaatori subkortikaalne kese paikneb: a) piklikajus b) sillas c) limbilises süsteemis d) talamuse lateraalsetes genikulaatkehades ja nelipealihase ülemistes kollikutes 44. Medulla oblongata keskpunkt visuaalne analüsaator asub ajukoores: a) kuklaluu b) parietaalne c) temporaalne d) eesmine 45. Silma võime eristada kahte helendavat punkti, mille projektsioonid langevad võrkkestale minutise nurga all, on nimetatakse: a) normaalne nägemisteravus b) silma murdumine c) presbüoopia d) astigmatism 46. Silma võime häälestuda objektide selgele nägemisele sõltuvalt nende kaugusest, neid nimetatakse: a) akommodatsiooniks b) nägemisteravus c) presbüoopia d) astigmatism 47. Silma akommodatsioon on tingitud peamiselt: muutusest: a) läätse kumerusest b) varraste arvust c) aktiivsete retseptorite arvust d) pupilli läbimõõdust An silma tundlikkuse suurenemine pimedas on seotud: a) jodopsiini lagunemisega b) jodopsiini sünteesiga c) rodopsiini sünteesiga d) rodopsiini lagunemisega
7 51. Silmade täielik kohanemine heledast ruumist lahkumisel pimedamaks toimub: a) 1-3 minutiga b) 4-5 minutiga c) minutiga d) minutiga 52. Silmade kohanemine pimedast ruumist lahkumisel ere valgus tekib: a) 1-3 minutiga b) 4-5 minutiga c) minutiga d) minutiga, mida nimetatakse: a) vaateväli b) vastuvõtuväli c) ruumilävi d) nägemisteravus Võrkkesta fotoretseptorite elektriline koguaktiivsus on nimetatakse: a) elektroretinogramm b) elektrokardiogramm c) elektroentsefalogramm d) kymogramm 57. Normaalne silmasisene rõhk inimesel on: a) 6-15 mm Hg. Art. b) mm Hg. Art. c) mm Hg. Art. d) mm Hg. Art. 58. Seniilne kaugnägelikkus, mis tekib inimestel aastate pärast, on: a) lühinägelikkus b) Presbüoopia c) Emmetroopia d) Astigmatism pulkade arv
8 60. Hüpermetroopia ja presbüoopia korral on põhifookus: a) võrkkesta taga b) võrkkesta ees c) võrkkestal 61. Müoopia (lühinägelikkuse) korral on põhifookus: a) võrkkesta ees b) võrkkesta ees. ) võrkkestal c) võrkkesta taga 62. Murdumise anomaalia, mille puhul valguskiired on fokuseeritud võrkkesta taha. on: a) Lühinägelikkus b) Emmetroopia c) Astigmatism d) Hüpermetroopia 63. Murdumisanomaalia, mille puhul valguskiired koonduvad võrkkesta ette, on: a) Emmetroopia b) Lühinägelikkus c) Hüpermetroopia d) Presbüoopia 64. Lühinägelikkus on korrigeeritud: a) silindriliste läätsedega b) astigmaatilise läätsega c) kaksikkumerate läätsedega d) kaksikkumerate läätsede hulka kuuluvad: a) trummikile, malleus, alasi, jalus b) Eustachia toru, vestibüül c) Corti elund, poolringikujulised kanalid Ninaneelus Thetympanic 68. selle maht on umbes: a) 1 cm 3 b) 2 cm 3 c) 3 cm 3 d) 4 cm 3
9 69. Kooklee on osa kõrvast: a) Väline b) Keskmine c) Sisemine 70. Spiraalne (Cortian) elund paikneb: a) keskmine skala b) scala vestibule c) scala tympani d) Trummiõõs 71. Endolümf asub: a) keskmises trepikojas b) vestibüüli trepikojas c) trummikiles d) trummiõõnes 72. Kuulmisanalüsaatori retseptorsektsioon sisaldab: a) karvarakke b) trummikile c) peamembraani d) ) sisemembraan : a) trummikile deformatsioonid b) karvarakkude deformatsioonid c) trummikile võnked d) perilümfi võnked 74. Kuulmisanalüsaatori retseptoris retseptori potentsiaali kujunemise ajal membraan: a) repolariseerib b) depolariseerib c) hüperpolariseerib 75. Kuulmisanalüsaatori subkortikaalne kese paikneb: a) pikliku medulla b) tindi c) limbilise süsteemi quadrigemina 76. Kuulmisanalüsaatori kortikaalne esitus paikneb: a) oimusagarates b) parietaalsagaras c) kuklaluu piirkonnas d) somatosensoorses ajukoores 77. Inimese helivibratsioonide tajumisala on vahemikus : a) Hz b) Hz c) Hz d) Hz
10 78. Kõnehelide võnkesagedus sekundis on vahemikus: a) Hz b) Hz c) Hz d) Hz 4. Maitseanalüsaatori füsioloogia 79. Retseptori potentsiaal maitsmispunga struktuurides esineb: a) maitserakus b) basaalrakkudes c) tugirakkudes d) maitsmiskanalis 80. Maitseretseptoritele viidatakse: a) kauge tüüp b) kontakttüüp 81. Mis tüüpi maitseanalüsaatori retseptorrakud need on ? a) Sekundaarne tunne b) Primaarne meel 82. Millised ioonid mängivad soolase maitse tundmise ajal peamist rolli retseptori potentsiaali tekitamisel? a) Ca2+ b) H+ c) Na+ d) Cl- 83. Millised ioonid mängivad peamist rolli retseptori potentsiaali tekkes haputunde ajal? a) Ca2+ b) H+ c) Na+ d) CI- 84. Millise maitsega toimub kohanemine kõige kiiremini? a) Magusaks b) Mõruks c) Glutamaadi maitseks d) Hapuks
11 5. Haistmisanalüsaatori füsioloogia 86. Märkige haistmisretseptori struktuur: a) Epiteelirakud b) Bipolaarsed neuronid c) Pseudounipolaarsed neuronid d) Haistmisbulbid 87. Mis tüüpi on haistmisretseptorid? a) interotseptiivne b) eksterotseptiivne c) propriotseptiivne 88. Mis tüüpi on haistmisretseptorid? a) Kontakt b) Kaug 89. Retseptor haistmisrakud liigitatakse: a) sekundaarseteks sensoorseteks b) primaarseteks sensoorseteks 90. Millises järjekorras saadetakse haistmisinformatsioon ajju? a) haistmisnärvid haistmissibulad haistmistrakt haistmiskolmnurk eesmine perforeeritud aine hipokampus b) haistmistrakti haistmissibulad haistmisnärvid haistmiskolmnurk eesmine perforeeritud aine hipokampus c) haistmisnärvikolmnurk - perforeeritud hippokampus ppokampus 91. Info töötlemise õige järjestus sisse haistmisanalüsaator: a) haistmisbulb eesaju b) haistmissibul keskaju eesaju c) haistmissibul talamuse eesaju d) haistmissibul ajukoore piirkonnad d) ajukoore somatosensoorset tsooni esindavad: ajukoore retseptorid3. ) A. Ruffini kehad b) V. Krause kolvid c) G. Meissneri kehad d) F. Merkeli kettad. 94. Naha külmaretseptoreid esindavad: a) A. Ruffini Sõnn b) W. Krause kolvid c) G. Meisneri Sõnn d) F. Merkeli kettad. 6. Temperatuurianalüsaatori füsioloogia
12 95. Nahas on sügavamalt lokaliseeritud: a) külmaretseptorid b) soojusretseptorid c) Pacini kehakesed 96. Seal on rohkem: a) soojusretseptoreid b) külma retseptoreid nahapinna ühiku kohta 97. Kortikaalne esitus temperatuurianalüsaator asub: a) pretsentraalne gyrus b) postcentral gyrus c) ajukoore kuklaluu d) ajukoore ajalises piirkonnas 98. Naha taktiilsed retseptorid on esindatud: . 99. Naha rõhu retseptorite hulka kuuluvad: a) A. Ruffini Sõnn b) G. Meissneri Sõnn c) A. Vateri Sõnn - F. Pacini d) Vabad närvilõpmed. 7. Puuteanalüsaatori füsioloogia 100. Minimaalset kaugust kahe punkti vahel, mille samaaegsel stimuleerimisel tekib kahe puudutuse tunnetus, nimetatakse: a) ruumilävi b) lävijõud c) ärrituslävi d) tundlikkuslävi 101. maksimaalsel ruumilisel lävel on: a) selg b) küünarvars c) käe tagakülg d) sõrm 102. Minimaalne ruumiline lävi on: a) sõrm b) küünarvars c) jalalaba plantaarne osa d) selg
13 8. Motoorse analüsaatori füsioloogia 103. Motoorse (propriotseptiivse) analüsaatori funktsioon on peamiselt omane lihastele: a) Süda b) Skelett c) Veresooned d) Siseorganid 104. Lihaste venitusretseptorid: a) Lihasvõllid b) ) Krause kolvid c) Merkeli kettad d) Meissneri kehad 105. Golgi kõõluselund paikneb: a) lihaste kõõlustes b) ekstrafusaalsete lihaskiudude hulgas c) intrafusaalsete kiudude distaalsetes lõikudes d) tuumas. intrafusaalsete kiudude kott 106. Intrafusaalsed lihaskiud täidavad järgmisi funktsioone: a) tagavad nõrga kontraktsiooni b ) tagavad lihaste spindli venitustundlikkuse c) lihaste lõdvestamise 9. Notsitseptiivse (valu) analüsaatori füsioloogia 107. kehakudede kahjustusest tuleneva valu tajumist nimetatakse: a) notsitseptsioon b) kiiritus c) analgeesia d) tajumine 108. Valuretseptorid: a) Meissneri kehad b) Krause kolvid c) vabad närvilõpmed d) Ruffini kehad
Analüsaatorite füsioloogia. Praegune kontrollkatse 1. Mõiste "analüsaator" tutvustas füsioloogias esmakordselt 1909. aastal N.E. Vvedensky A.A. Ukhtomsky I.P. Pavlov C. Sherrington 2. Valige kõige täpsem
ANDURID. RECEPTORID. TEABE KODEERIMISE PÕHIMÕTTED. SENSORSED RETSEPTORID Sensoorsed retseptorid on spetsiifilised rakud, mis on häälestatud tajuma erinevaid välis- ja sisekeskkonna stiimuleid.
Keha sensoorsete süsteemide arendamine Sensoorsed süsteemid (analüsaatorid) on ühtsed teabe analüüsimise süsteemid, mis koosnevad 3 osakonnast: perifeerne, juhtiv ja tsentraalne. Osakonnad (lingid) Välisseadmed
8. klass Teema: Analüsaatorid ehk andursüsteemid Andurisüsteemide üldised omadused. Nende struktuur, funktsioonid. Sensoorsete süsteemide põhilised füsioloogilised omadused. visuaalne analüsaator. Silma struktuur. murduv
8. klass Bioloogia profiil Teema: Meeleelundid Ülesanne 1 Sensoororganid Nägemisretseptorid asuvad silma kestas, mida nimetatakse ... [Retinal Iris Vascular Cornea] 2. ülesanne Sensoorsed organid
Analüsaatorid ja meeleorganid Analüsaator sisaldab 3 komponenti: Perifeerne osa (retseptorid, sensoorne organ) Juhtosa (närvikiud) Keskosa (ajukoore tsoon) Tajub
Analüsaator (kreeka keeles analüüs, lagunemine, tükeldamine) on närvistruktuuride kogum, mis tajub ja analüüsib erinevaid väliseid ja sisemisi stiimuleid. Selle termini pakkus välja I. P. Pavlov 1909. aastal.
Analüsaatorid, meeleelundid ja nende tähendus Analüsaatorid. Kõik elusorganismid, sealhulgas inimesed, vajavad teavet keskkonna kohta. Selle võimaluse pakuvad neile sensoorne (tundlik)
Biofüüsikalised protsessid välis-, kesk- ja sisekõrvas. Kuulmissensoorne süsteem sisaldab: Väliskõrva ehitust. Väliskõrva funktsioonid. Kuulmistaju orientatsioon. Keskkõrv (trummiks
Bioloogia test Analüsaatorid Sensoororganid 8. hinne 1 valik 1. Sensoorsete organite ülesanne on muundada välisärrituse energia ärritusele ligipääsetavasse vormi A. Retseptorid B. Seljaaju
Rahvaste sõpruse ülikool Venemaa Meditsiiniinstituut Inimese anatoomia osakond Eriala: õendusdotsent Gurova O.A. MEELEELUNDID Loengukava: 1. Meeleelundite ehitusmustrid
Tundlikkuse tüübid (vastuvõtt) eksterotseptiivne üldine (somatosensoorne) - taktiilne, valu, temperatuuri eriline visuaalne kuulmine haistmine maitsmis-gravitatsiooniline (tasakaal) interotseptiivne
LÕPUTESTID rubriigis ANALÜÜSIJATE FÜSIOLOOGIA (SENSOORSÜSTEEMID) Vali üks õige vastus 1. Retseptorite tundlikkuse vähenemist nimetatakse: a) erutuvuseks b) spetsiifilisuseks
MEELEELUNDID Nägemisorganid Meeleelundid (analüsaatorid) Anatoomilised moodustised (seadmed) (i) välismõju energia tajumine, (ii) närviimpulssiks muutmine ja (iii) edasikandmine
VENEMAA FÖDERATSIOONI HARIDUSMINISTEERIUM IRKUTSK RIIKLIKÜLIKOOL Bioloogia ja mullateaduse teaduskond füsioloogia ja psühhofüsioloogia osakond KINNITUD Õppemeetodite teaduskonna esimehe poolt 2004: PROGRAMM
National Pharmaceutical University Inimese füsioloogia ja anatoomia osakond Visuaalne analüsaator. Analüsaatorite vanuseomadused Shatalova O.M. Plaan 1. Sensoorsete süsteemide ehituse üldpõhimõtted.
TEEMA "Analüsaatorid" 1. Haistmisanalüsaatori alglüliks loetakse 1) närvid ja närviteed 2) keelel asuvad retseptorid 3) ajukoore neuronid 4) tundlikud
304-Rühm: Fattoeva Zarina. Kontrollinud: Rakhmatova N.B. Samarkand - 2016 FUNKTSIOONIAALSETE SÜSTEEMIDE TEOORIA Petr Kuzmich Anokhin (1898-1974) Funktsionaalse süsteemi dünaamiline isereguleeruv organisatsioon, kõik
Loeng 6. Mentaalsed kognitiivsed aistingud ja tajuprotsessid: 6.2 Sensatsioonide mõiste A.V. Petrovski sõnul on aistingud otseselt mõjutavate objektide ja nähtuste individuaalsete omaduste peegeldus
Küsimuste loetelu lõplikuks kontrolliks Kesknärvisüsteem. 1. Kesknärvisüsteemi areng embrüogeneesis. Närvisüsteemi kujunemise peamised etapid fülogeneesis. 2. Aju areng
LÕPUTUND JAGUS „NÄRVISÜSTEEMI KONKREETSED FÜSIOLOOGIA. SENSOORSÜSTEEMIDE FÜSIOLOOGIA» Põhiküsimused: 1. Seljaaju. Seljaaju funktsioonid. Põhilised seljaaju refleksid. Kahjustuse tagajärjed
1 1.7. Inimese analüsaatorid 1.7.1. analüsaatori seade. Visuaalne analüsaator Inimese keskkonnatingimuste ja sisekeskkonna seisundi muutusi tajub närvisüsteem, mis reguleerib.
TÖÖPROGRAMMI "NEUROFÜSIOLOOGIA" MÄRKUS Rakendatud koolitusvaldkonna spetsialisti koolitamise õppekava põhiosas (spetsialist) GEF 37.05.01 / kliiniline psühholoogia
NÄRVISÜSTEEM. ANDURID. 1. Neuron: definitsioon, osad, morfoloogiline klassifikatsioon, struktuur, topograafia, 2. Lihtsa ja keeruka refleksikaare ehitus 3. Kesknärvisüsteemi areng
Sensoorne süsteem Vali üks õige vastus 001. Võrkkesta areneb 1) silmakupa sisekihist 2) silmakupa väliskihist 3) silmapõiekese ees paiknevast ektodermist.
Teema: NÄRVISÜSTEEM (6 tundi). Üldine ülevaade närvisüsteemist. Närvisüsteemi ehitus ja talitlus. Klassifikatsioon topograafiliste ja funktsionaalsete tunnuste järgi. Neuron põhiline struktuurne-funktsionaalne
TESTID Sensoorsete süsteemide üldfüsioloogia Nägemise füsioloogia Tasakaalustaju ja kuulmise füsioloogia Somatovistseraalne tundlikkus, valu Loeng 1 Sensoorsete süsteemide üldfüsioloogia 1. *Milline nähtus
Jooksvad kontrolltestid teemal Närvisüsteemi spetsiifiline füsioloogia 1. Millistes seljaaju sarvedes paiknevad alfamotoorsete neuronite kehad? a) Tagumises b) Külgmises c) Eesmises 2. Seljaajus sulge
Ligikaudsed ülesanded bioloogias P4 8. klass 1. Millises ajukooresagaras asub kuulmistsoon: A) frontaalne B) kuklaluu C) parietaalne D) ajaline 2. Mitu aksonit võib närvirakul olla: A)
STRELNIKOVA BIOLOOGIA JA SILMADE ARENG VICTORIA VIKTOROVNA, SBEI IRO KK (ARMAVIR IRINSEYESI KIRI) HARIDUSTEGEVUSE TEADUSLIKU JA METOODILISE TOETUSE OSAKONNA METOODIKA
Inimese analüsaatorite omadused Inimese analüsaator on kesknärvisüsteemi alamsüsteem, mis võimaldab informatsiooni vastuvõtmist ja esmast analüüsi. Analüsaatori retseptori perifeerne osa, keskne
Optiliste kujutiste geomeetriline teooria Kui punktist A mittehomogeenses keskkonnas peegeldumise, murdumise või paindumise tulemusena väljuv valguskiir koondub punktis A, siis A
1 - "KINNITUD" Normaalse füsioloogia osakonna juhataja, meditsiiniteaduste doktor, professor S.V. Klauceki protokoll 1, 29. august 2014
Vestibulaarsed ja kinesteetilised analüsaatorid 1. Vestibulaarse analüsaatori ülesehitus 2. Kinesteetilise analüsaatori ülesehitus 3. Sisemised (vistseraalsed) analüsaatorid Küsimus_1 Vestibulaarse analüsaatori korraldus
POOLKERA KORREKSI FUNKTSIONAALNE ORGANISATSIOON 1 Aju üldine korraldus 2 Aju integreeriva töö struktuurne ja funktsionaalne mudel (Luria A.R.) 3 Teletsefaloni moodustavad kaks poolkera, mis
KUULMISANALÜÜSER Muusika üldise toimemehhanismi mõistmine inimkehale on võimatu ilma kuulmisanalüsaatori ehituse ja tööpõhimõtete tundmiseta. Kuulmisanalüsaator on loodud tajuma
RF ORAGOAAIA JA TEADUSTE MINISTER Murmanski Riiklik Humanitaarülikool (FOU PO "MU")
ANALÜÜSID ANALÜÜSIJATE ÜLDOMADUSED 1. Retseptoris on kodeeritud stiimuli tugevus: 1. retseptori potentsiaali esinemise sagedus 2. retseptori potentsiaali amplituud 2. retseptorid spetsialiseerunud
MATERJALID testimiseks ettevalmistamiseks 8. klassis Õpetaja: Kuturova Galina Aleksejevna TEEMA Rubriik "Närvisüsteem" Rubriik "Visuaalne analüsaator" TEADA / OLLA Tähendus, struktuur ja toimimine
3 Sisukord Sissejuhatus. 4 1. jagu. Närvisüsteem ja analüsaatorid.5 1.1. Närvisüsteemi funktsioonid ja struktuur 6 1.1.1. Kesknärvisüsteem.11 1.1.2. Autonoomne närvisüsteem 15 1.2. Tähendus ja
Füsioloogia anatoomia alustega Auditoorsed ja vestibulaarsed analüsaatorid Ph.D. Assoc. Kuchuk A.V. Kuulmisanalüsaator Piisava stiimuli mehaaniline laine vahemikus 20 20000 Hz Mehaanilise laine parameetrid
ANDURID: Kuulmis- ja tasakaaluorgan Haistmiselund Maitseelund Nahk Vestibulokohleaarne elund (kuulmis- ja tasakaaluorgan) See jaguneb kolmeks anatoomiliselt ja funktsionaalselt ühendatud osaks:
Vene Föderatsiooni transpordiministeerium Föderaalne riigieelarveline kõrgharidusasutus "VENEMAA TRANSPORDIÜLIKOOL (MIIT)" psühholoogia, sotsioloogia osakond,
TÖÖTERVISHOIU FÜSIOLOOGILISED ALUSED Õpik Peterburi 2006 Vene Föderatsiooni Haridus- ja Teadusministeerium Föderaalne Haridusagentuur ST PETERBURGI RIIKLIKÜLIKOOL
Teema: Kesknärvisüsteem. Seljaaju ja aju. Perifeerne närvisüsteem. 1-variant 1. Ajutüvi on: 1) sild, piklik medulla 2) piklik medulla 3) keskaju, sild
A. Baitursynovi nimeline Kostanay Riiklik Ülikool Lühiandmed nägemisorgani füsioloogia kohta Dotsent Baikenov M.T. Loomade visuaalse analüsaatori põhifunktsioon on valguse tajumine,
Närvilõpmed, klassifikatsioon Lõppseadmed (interneuronaalsed sünapsid) Efektornärvilõpmed (efektorid, neuroorganite sünapsid) Sensoorsed (retseptorid) närvilõpmed Sünapsid dendriidid
KUULMISE PÕHIOMADUSED Inimese kuulmisorgan on omamoodi helivastuvõtja, mis erineb järsult inimese loodud helivastuvõtjatest. Inimkõrval on sagedusanalüsaatori omadused,
UZBEKISTANI VABARIIGI TERVISEMINEERIUM SAMARKANDI MEDITSIINI INSTITUUT KOKKUVÕTE TEEMA: SELJAAJU Lõpetanud: Vohidov U. SAMARKAND-2016 SELJAAJU Närvisüsteemi tähtsus Närvisüsteem
NAHAANALÜÜSID 1. Naha ehitus ja retseptorite asukoht 2. Puuteanalüsaatori ehitus ja funktsioonid 3. Temperatuurianalüsaatori ehitus ja funktsioonid Küsimus_1 Naha ehitus ja retseptorite asukoht
Silm ja selle funktsioonid Loeng 1. Silma ehitus. Majutus. binokulaarne nägemine. 2. Silma optilise süsteemi puudused. 3. Vaatenurk. Resolutsioon. Nägemisteravus. 4. Akustiline biomehaanika
Hindamisvahendite fond eriala üliõpilaste keskastme atesteerimise läbiviimiseks (moodul): Üldinfo 1. Loodusteaduste osakond 2. Koolituse suund 06.03.01 Bioloogia, profiil Üldine
Teoreetilise osa küsimused NEUROLOOGIA (KNS) LÕPUTUND 1. Närvisüsteemi filo- ja ontogenees. 2. Närvisüsteemi osakonnad ja nende tähendus. 3. Neuron on närvisüsteemi struktuurne ja funktsionaalne üksus.
13. loeng Suu- või suuanalüsaatori mõisted, roll toitainete aprobeerimisel. Maitse ja lõhn
Geomeetrilise optika alused. Inimese nägemise aparatuur Plaan 1. Geomeetrilise optika põhimõisted. 2. Silma valgust juhtivad ja valgust tajuvad süsteemid. 3. Nägemise puudumine. Valgus on elektromagnetiline
MATERJALID Bioloogia klassi 8.1 ettevalmistamiseks 4. moodul Õpetaja: Z.Yu. Sobolev Sektsioon / Teema Teadma Oskama tajuorganeid Nägemisaparaadi ehitus Kuulmisorgani ehitus ja vestibulaaraparaat Alused
TEEMA "Närvisüsteem" 1. Millist funktsiooni täidab närvirakk inimese ja looma kehas 1) motoorne 2) kaitsev 3) ainete transport 4) ergastuse läbiviimine 2. Millises ajuosas see asub
EKSKÜSIMUSTE LOETELU Neuroanatoomia kui teadus 1. Kesknärvisüsteemi morfoloogilise ja funktsionaalse korralduse vaadete ja õpetuste kujunemislugu (R. Descartes, F. Gall, V. Betz jt).
Perekonnanimi Kood Nimi Piirkond Töökoha kood Punktid kokku ÜLESANNE (demoversioon) piirkondadevahelise bioloogiaolümpiaadi "ALFA" praktiline ringkäik, 2014-2015 konto. aasta, 9. klass Demonstratsioon
Tunded BIOLOOGIA INIMTUNNED 1. peatükk: Meie tunded Miks me vajame oma tundeid? Kõik organismid tunnevad oma ümbrust, kuid loomadel ja inimestel on välja kujunenud mõned väga keerulised sensoorsed süsteemid,
Distsipliini (mooduli) "Normaalne füsioloogia" tööprogrammi annotatsioon suunal 14.03.02 Tuumafüüsika ja -tehnoloogia (profiil Inimese ja keskkonna kiirgusohutus) 1. Eesmärgid ja eesmärgid
1. loeng SENSORSÜSTEEMIDE ÜLDFÜSIOLOOGIA Taju objektiivne ja subjektiivne pool Sensoorsete süsteemide spetsiifilisus Konkreetsete energiate seadus Sensoorse süsteemi ehitus Sensoorsete organisatoorsete süsteemide põhimõtted.
Sissepääsu kontrolltöö bioloogiaklassis 9 1 variant 1. Veri viitab koe tüübile: A) sidekoe B) närviline C) epiteel D) lihaseline 2. Vaagna lihased hõlmavad A) tuharalihaseid B) gastrocnemius
Tunni teema: Analüsaatorite tundlikkus. Analüsaatorite koostoime. Bioloogiaõpetaja Burmistrova Inna Evgenievna tund Tunni eesmärgid: jätkata meeleelundite mõistete kujundamist; korda ja võta kokku
Fundamentals of psychophysiology., M. INFRA-M, 1998, lk 57-72, 2. peatükk Ed. Yu.I. Aleksandrov
2.1. Silma optilise aparaadi ehitus ja funktsioonid
Silmamuna on kerakujuline, mis muudab vaadeldavale objektile sihtimiseks pööramise lihtsamaks ja tagab pildi hea teravustamise kogu silma valgustundlikule kestale – võrkkestale. Teel võrkkestale läbivad valguskiired mitut läbipaistvat keskkonda – sarvkesta, läätse ja klaaskeha. Sarvkesta ja vähesel määral ka läätse teatud kumerus ja murdumisnäitaja määravad valguskiirte murdumise silma sees. Võrkkestale saadakse kujutis, mis on järsult vähendatud ja pööratud tagurpidi ja paremalt vasakule (joonis 4.1 a). Iga optilise süsteemi murdumisvõimet väljendatakse dioptrites (D). Üks diopter võrdub 100 cm fookuskaugusega läätse murdumisvõimega.Terve silma murdumisvõime on kaugel asuvate objektide vaatamisel 59D ja lähedal asuvate objektide vaatamisel 70,5D.
Riis. 4.1.
2.2. Majutus
Akommodatsioon on silma kohanemine erinevatel kaugustel asuvate objektide selge nägemisega (sarnaselt teravustamisega fotograafias). Objekti selgeks nägemiseks on vajalik, et selle kujutis oleks fokuseeritud võrkkestale (joonis 4.1 b). Akommodatsioonis mängib põhirolli läätse kumeruse muutus, s.o. selle murdumisvõime. Lähedasi objekte vaadates muutub objektiiv kumeramaks. Akommodatsioonimehhanism on läätse kumerust muutvate lihaste kokkutõmbumine.
2.3. Silma murdumishäired
Silma kaks peamist murdumisviga on lühinägelikkus (lühinägelikkus) ja kaugnägelikkus (hüpermetroopia). Need kõrvalekalded ei ole tingitud silma murdumiskeskkonna puudulikkusest, vaid silmamuna pikkuse muutusest (joonis 4.1 c, d). Kui silma pikitelg on liiga pikk (joonis 4.1 c), siis kaugemal asuva objekti kiired ei keskendu mitte võrkkestale, vaid selle ette, klaaskehasse. Sellist silma nimetatakse lühinägelikuks. Et näha selgelt kaugusesse, peab lähinägija asetama oma silmade ette nõgusad prillid, mis suruvad teravustatud pildi võrkkestale (joonis 4.1 e). Seevastu kaugnägevas silmas (joonis 4.1 d) pikitelg lüheneb ja seetõttu on kauge objekti kiired fokuseeritud võrkkesta taha. Seda puudust saab kompenseerida läätse kumeruse suurenemisega. . Lähedasi objekte vaadates on aga kaugelenägevate inimeste kohanemispingutused ebapiisavad. Seetõttu peavad nad lugemiseks kandma kaksikkumerate läätsedega prille, mis suurendavad valguse murdumist (joonis 4.1 e).
2.4. Pupill ja pupillide refleks
Pupill on auk iirise keskel, mille kaudu valgus siseneb silma. See suurendab võrkkesta kujutise selgust, suurendades silma teravussügavust ja kõrvaldades sfäärilise aberratsiooni. Laienduse korral kitseneb valguses olev pupill kiiresti ("pupillirefleks"), mis reguleerib silma siseneva valguse voolu. Niisiis, eredas valguses on pupilli läbimõõt 1,8 mm, keskmise päevavalguse korral laieneb see 2,4 mm-ni ja pimedas kuni 7,5 mm. See halvendab võrkkesta pildikvaliteeti, kuid suurendab nägemise absoluutset tundlikkust. Õpilase reaktsioonil valgustuse muutustele on adaptiivne iseloom, kuna see stabiliseerib võrkkesta valgustust väikeses vahemikus. Tervetel inimestel on mõlema silma pupillid sama läbimõõduga. Kui üks silm on valgustatud, kitseneb ka teise silmapupill; sellist reaktsiooni nimetatakse sõbralikuks.
2.5. Võrkkesta ehitus ja funktsioonid
Võrkkesta on silma sisemine valgustundlik membraan. Sellel on keeruline mitmekihiline struktuur (joonis 4.2). On kahte tüüpi fotoretseptoreid (vardad ja koonused) ja mitut tüüpi närvirakke. Fotoretseptorite ergastamine aktiveerib võrkkesta esimese närviraku - bipolaarse neuroni. Bipolaarsete neuronite ergastamine aktiveerib võrkkesta ganglionrakud, mis edastavad oma impulsid subkortikaalsetesse nägemiskeskustesse. Horisontaalsed ja amakriinsed rakud osalevad ka võrkkesta teabe edastamise ja töötlemise protsessides. Kõik need võrkkesta neuronid koos oma protsessidega moodustavad silma närviaparaadi, mis on seotud visuaalse informatsiooni analüüsi ja töötlemisega. Seetõttu nimetatakse võrkkesta perifeeriasse paigutatud aju osaks.
2.6. Võrkkesta kihtide struktuur ja funktsioonid
Rakud pigmendi epiteel moodustavad võrkkesta välimise, valgusest kõige kaugema kihi. Need sisaldavad melanosoome, mis annavad neile musta värvi. Pigment neelab liigset valgust, takistades selle peegeldumist ja hajumist, mis aitab kaasa pildi selgusele võrkkestal. Pigmendiepiteel mängib otsustavat rolli fotoretseptorite visuaalse lilla regenereerimisel pärast selle värvimuutust, visuaalsete rakkude välissegmentide pidevas uuendamises, retseptorite kaitsmisel valguskahjustuste eest, samuti hapniku ülekandmisel ja neile toitaineid.
Fotoretseptorid. Visuaalsete retseptorite kiht: vardad ja koonused külgnevad seestpoolt pigmendiepiteeli kihiga. Iga inimese võrkkest sisaldab 6-7 miljonit koonust ja 110-125 miljonit varrast. Need on võrkkestas jaotunud ebaühtlaselt. Võrkkesta keskne fovea - fovea (fovea centralis) sisaldab ainult koonuseid. Võrkkesta perifeeria suunas koonuste arv väheneb ja varraste arv suureneb, nii et kaugemal on ainult vardad. Koonused toimivad kõrge valgustuse tingimustes, need tagavad päeva- ja värvinägemise; valgustundlikumad vardad vastutavad hämara nägemise eest.
Värvi tajutakse kõige paremini, kui valgus tabab võrkkesta fovea, mis sisaldab peaaegu eranditult käbisid. Siin on suurim nägemisteravus. Võrkkesta keskpunktist eemaldudes vähenevad järk-järgult värvitaju ja ruumiline eraldusvõime. Võrkkesta perifeeria, mis sisaldab ainult vardaid, ei taju värve. Teisest küljest on võrkkesta koonusaparaadi valgustundlikkus mitu korda väiksem kui vardaaparaadil. Seetõttu ei erista me hämaras koonuse nägemise järsu vähenemise ja perifeerse varraste nägemise ülekaalu tõttu värvi ("kõik kassid on öösel hallid").
visuaalsed pigmendid. Inimese võrkkesta vardad sisaldavad pigmenti rodopsiini ehk visuaalselt lillat, mille maksimaalne neeldumisspekter jääb 500 nanomeetri (nm) vahemikku. Kolme tüüpi koonuste (sinise-, rohelise- ja punase-tundlikud) välimised segmendid sisaldavad kolme tüüpi visuaalseid pigmente, mille neeldumisspektri maksimumid on sinises (420 nm), rohelises (531 nm) ja punases ( 558 nm) spektri piirkonnad. Punast koonuse pigmenti nimetatakse jodopsiiniks. Visuaalse pigmendi molekul koosneb valguosast (opsiin) ja kromofooriosast (võrkkesta ehk A-vitamiini aldehüüd). Võrkkesta allikaks kehas on karotenoidid; nende puudusega hämaras nägemine ("ööpimedus") on halvenenud.
2.7. võrkkesta neuronid
Võrkkesta fotoretseptorid on sünaptiliselt ühendatud bipolaarsete närvirakkudega (vt joonis 4.2). Valguse toimel väheneb vahendaja vabanemine fotoretseptorist, mis hüperpolariseerib bipolaarse raku membraani. Sellest edastatakse närvisignaal ganglionrakkudesse, mille aksoniteks on nägemisnärvi kiud.

Riis. 4.2. Võrkkesta struktuuri skeem:
1 - pulgad; 2 - koonused; 3 - horisontaalne rakk; 4 - bipolaarsed rakud; 5 - amakriinrakud; 6 - ganglionrakud; 7 - nägemisnärvi kiud
Iga 130 miljoni fotoretseptori raku kohta on ainult 1 250 000 võrkkesta ganglionrakku. See tähendab, et paljude fotoretseptorite impulsid koonduvad (koonduvad) bipolaarsete neuronite kaudu ühte ganglionrakku. Ühe ganglionrakuga ühendatud fotoretseptorid moodustavad selle vastuvõtuvälja [Huebel, 1990; Physiol. nägemus, 1992]. Seega võtab iga ganglionrakk kokku paljudes fotoretseptorites esineva ergastuse. See suurendab võrkkesta valgustundlikkust, kuid halvendab selle ruumilist eraldusvõimet. Ainult võrkkesta keskel (fovea piirkonnas) on iga koonus ühendatud ühe bipolaarse rakuga, mis omakorda on ühendatud ühe ganglionrakuga. See tagab võrkkesta keskkoha kõrge ruumilise eraldusvõime, kuid vähendab järsult selle valgustundlikkust.
Võrkkesta naaberneuronite interaktsiooni tagavad horisontaalsed ja amakriinsed rakud, mille käigus levivad signaalid, mis muudavad sünaptilist ülekannet fotoretseptorite ja bipolaarsete (horisontaalsed rakud) ning bipolaarsete ja ganglionrakkude (amakriinide) vahel. Amakriinrakud inhibeerivad külgnevaid ganglionrakke lateraalselt. Võrkkestale tulevad ka tsentrifugaalsed ehk eferentsed närvikiud, mis toovad ajust sinna signaale. Need impulsid reguleerivad ergastuse juhtivust võrkkesta bipolaarsete ja ganglionrakkude vahel.
2.8. Närvide rajad ja ühendused nägemissüsteemis
Võrkkestast liigub visuaalne informatsioon mööda nägemisnärvi kiude ajju. Kahe silma närvid kohtuvad aju põhjas, kus osa kiududest liigub vastasküljele (optiline kiasm või kiasm). See annab igale ajupoolkerale informatsiooni mõlemast silmast: parema poolkera kuklasagara saab signaale iga võrkkesta parempoolselt poolelt ja vasak poolkera saab signaale kummagi võrkkesta vasakust poolest (joonis 4.3).
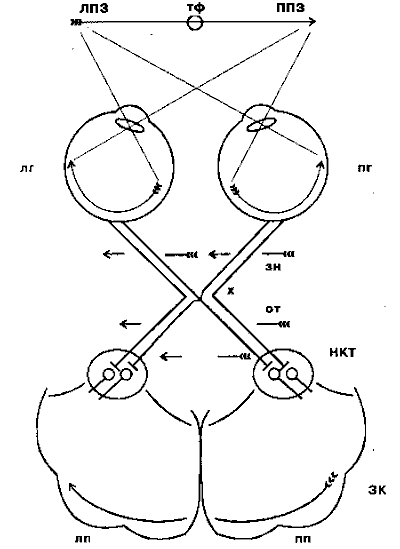
Riis. 4.3. Visuaalsete radade skeem võrkkestast primaarse nägemiskooreni:
LPZ - vasak vaateväli; RPV - parem vaateväli; tf - pilgu fikseerimise punkt; lg - vasak silm; pg - parem silm; zn - nägemisnärv; x - optiline kiasm või kiasm; alates - optiline tee; torud - väline geniculate keha; ZK - visuaalne ajukoor; lp - vasak poolkera; pp - parem ajupoolkera
Pärast kiasmi nimetatakse nägemisnärve optilisteks traktideks ja enamik nende kiude jõuab subkortikaalsesse nägemiskeskusesse - lateraalsesse geniculate body (NKT). Siit sisenevad visuaalsed signaalid visuaalse ajukoore esmasesse projektsioonipiirkonda (vöötkoore ehk Brodmanni järgi väli 17). Nägelik ajukoor koosneb mitmest väljast, millest igaüks täidab oma spetsiifilisi funktsioone, võttes vastu nii otseseid kui kaudseid signaale võrkkestalt ja säilitades üldiselt selle topoloogia ehk retinotoopia (võrkkesta naaberpiirkondade signaalid sisenevad ajukoore naaberpiirkondadesse ).
2.9. Visuaalse süsteemi keskuste elektriline aktiivsus
Valguse toimel retseptorites ja seejärel võrkkesta neuronites tekivad elektrilised potentsiaalid, mis peegeldavad toimiva stiimuli parameetreid (joonis 4.4a, a). Võrkkesta kogu elektrilist reaktsiooni valgusele nimetatakse elektroretinogrammiks (ERG).

Riis. 4.4. Nägemiskoore elektroretinogramm (a) ja valguse esilekutsutud potentsiaal (EP) (b):
a, b, c, d kohta (a) - ERG lained; nooled näitavad valguse sisselülitamise hetki. R 1 - R 5 - positiivsed EP lained, N 1 - N 5 - negatiivsed EP lained (b)
Seda saab salvestada tervest silmast: üks elektrood asetatakse sarvkesta pinnale ja teine silma lähedal olevale näonahale (või kõrvapulgale). ERG peegeldab hästi valgusstiimuli intensiivsust, värvi, suurust ja kestust. Kuna ERG-s kajastub peaaegu kõigi võrkkesta rakkude (v.a ganglionrakud) aktiivsus, kasutatakse seda näitajat laialdaselt võrkkesta töö analüüsimisel ja haiguste diagnoosimisel.
Võrkkesta ganglionrakkude ergastumine toob kaasa asjaolu, et elektrilised impulsid sööstavad mööda nende aksoneid (nägemisnärvi kiude) ajju. Võrkkesta ganglionrakk on võrkkesta esimene "klassikalist" tüüpi neuron, mis genereerib levivaid impulsse. Kirjeldatud on kolme peamist ganglionrakkude tüüpi: reageerimine valguse sisselülitamisele (sisse - reaktsioon), selle väljalülitamine (väljas - reaktsioon) ja mõlemad (sisse-välja - reaktsioon). Võrkkesta keskosas on ganglionrakkude vastuvõtlikud väljad väikesed, võrkkesta perifeerias aga palju suurema läbimõõduga. Lähedal asuvate ganglionrakkude samaaegne ergastamine põhjustab nende vastastikust inhibeerimist: iga raku vastused muutuvad väiksemaks kui ühe stimulatsiooni korral. See toime põhineb lateraalsel või lateraalsel inhibeerimisel (vt ptk 3). Võrkkesta ganglionrakkude retseptiivsed väljad toodavad oma ümara kuju tõttu võrkkesta kujutise nn punkt-punkti kirjelduse: seda kuvab väga õhuke diskreetne mosaiik, mis koosneb ergastatud neuronitest.
Subkortikaalse nägemiskeskuse neuronid on erutatud, kui nad saavad võrkkestalt impulsse mööda nägemisnärvi kiude. Nende neuronite vastuvõtuväljad on samuti ümmargused, kuid väiksemad kui võrkkestas. Nende poolt valgussähvatuse tekitatud impulsspursked on lühemad kui võrkkestas. LNT tasemel toimub võrkkestast tulevate aferentsete signaalide interaktsioon visuaalse ajukoore eferentsete signaalidega, samuti kuulmis- ja muude sensoorsete süsteemide retikulaarformatsiooniga. See interaktsioon aitab eraldada signaali kõige olulisemad komponendid ja võib-olla osaleb selektiivse visuaalse tähelepanu korraldamises (vt 9. peatükk).
NKT neuronite impulsslahendused piki nende aksoneid sisenevad ajupoolkerade kuklaosasse, kus asub visuaalse ajukoore (vöötkoore) esmane projektsiooniala. Siin on primaatide ja inimeste puhul teabe töötlemine palju spetsialiseerunud ja keerukam kui võrkkesta ja LNT puhul. Nägemiskoore neuronitel ei ole ümmargused, vaid piklikud (horisontaalselt, vertikaalselt või diagonaalselt) väikesed vastuvõtuväljad (joon. 4.5) [Huebel, 1990].

Riis. 4.5. Kassi aju nägemiskoores oleva neuroni vastuvõtuväli (A) ja selle neuroni reaktsioonid vastuvõtuväljas (B) vilkuvatele erineva orientatsiooniga valgusribadele. A - plussidega on märgitud vastuvõtuvälja ergastav tsoon ja miinustega kaks külgmist inhibeerivat tsooni. B - on näha, et see neuron reageerib kõige tugevamalt vertikaalsele ja sellele lähedasele orientatsioonile
Tänu sellele suudavad nad pildilt valida üksikuid ühe või teise orientatsiooni ja asukohaga joonte fragmente ning neile valikuliselt reageerida. (orientatsioonidetektorid). Nägemiskoore igas väikeses piirkonnas, piki selle sügavust, on neuronid koondunud vaateväljas sama orientatsiooni ja vastuvõtuväljade lokaliseerimisega. Need moodustavad orientatsiooni veerg neuronid, mis läbivad vertikaalselt kõiki ajukoore kihte. Veerg on näide sarnast funktsiooni täitvate kortikaalsete neuronite funktsionaalsest ühendusest. Rühm naaberorientatsiooni veerge, mille neuronitel on kattuvad vastuvõtuväljad, kuid erinevad eelistatud orientatsioonid, moodustab nn superkolonni. Nagu näitavad viimaste aastate uuringud, võib nägemiskoores üksteisest kaugel olevate neuronite funktsionaalne ühinemine toimuda ka nende väljavoolude sünkroniseerimise tõttu. Viimasel ajal on nägemiskoorest leitud rist- ja nurkkujude suhtes selektiivse tundlikkusega neuroneid, mis kuuluvad 2. järku detektoritesse. Nii hakkas "nišš" pildi ruumilisi tunnuseid kirjeldavate lihtsate orientatsioonidetektorite ja ajalises ajukoores leiduvate kõrgemat järku (näo)detektorite vahel täituma.
Viimastel aastatel on hästi uuritud neuronite nn ruumisageduslikku häälestamist nägemiskoores [Glezer, 1985; Physiol. nägemus, 1992]. See seisneb selles, et paljud neuronid reageerivad valikuliselt teatud laiusega heledate ja tumedate triipude võrele, mis on ilmunud nende vastuvõtuvälja. Niisiis, on rakke, mis on tundlikud väikeste triipude võre suhtes, st. kõrgele ruumilisele sagedusele. Leiti erinevate ruumiliste sageduste suhtes tundlikud rakud. Arvatakse, et see omadus annab visuaalsele süsteemile võimaluse eristada pildilt erineva tekstuuriga alasid [Glezer, 1985].
Paljud visuaalse ajukoore neuronid reageerivad valikuliselt teatud liikumissuundadele (suunadetektorid) või mõnele värvile (värviga vastandlikud neuronid) ja mõned neuronid reageerivad kõige paremini objekti suhtelisele kaugusele silmadest. Infot visuaalsete objektide erinevate tunnuste (kuju, värv, liikumine) kohta töödeldakse paralleelselt nägemiskoore erinevates osades.
Et hinnata signaali edastamist visuaalse süsteemi erinevatel tasanditel, kogusumma registreerimine esile kutsutud potentsiaalid(VP), mida inimestel saab üheaegselt eemaldada võrkkestast ja nägemiskoorest (vt joonis 4.4 b). Välk-indutseeritud võrkkesta vastuse (ERG) ja kortikaalse EP võrdlus võimaldab hinnata projektsiooni visuaalse raja tööd ja teha kindlaks patoloogilise protsessi lokaliseerimine visuaalses süsteemis.
2.10. Valgustundlikkus
Absoluutne nägemistundlikkus. Visuaalse aistingu tekkimiseks peab valgusel olema teatud minimaalne (lävi)energia. Minimaalne valguskvantide arv, mis on vajalik valgustunde tekkimiseks pimedas, jääb vahemikku 8–47. Ühte pulka saab ergutada vaid 1 valguskvant. Seega on võrkkesta retseptorite tundlikkus valguse tajumiseks kõige soodsamates tingimustes piiriks. Võrkkesta üksikud vardad ja koonused erinevad veidi valgustundlikkuse poolest. Ühele ganglionrakku signaale saatvate fotoretseptorite arv on võrkkesta keskel ja perifeerias aga erinev. Koonuste arv võrkkesta keskel asuvas vastuvõtuväljas on umbes 100 korda väiksem kui võrkkesta perifeeria vastuvõtuväljas asuvate varraste arv. Vastavalt sellele on varraste süsteemi tundlikkus 100 korda suurem kui koonussüsteemil.
2.11. Visuaalne kohanemine
Pimedusest valgusele üleminekul tekib ajutine pimedus ja seejärel väheneb järk-järgult silma tundlikkus. Seda visuaalse süsteemi kohanemist ereda valguse tingimustega nimetatakse valguse kohanemiseks. Vastupidine nähtus (pimeda kohanemine) ilmneb siis, kui inimene liigub valgusküllasest ruumist peaaegu valgustamata ruumi. Algul ei näe ta fotoretseptorite ja visuaalsete neuronite vähenenud erutuvuse tõttu peaaegu midagi. Järk-järgult hakkavad ilmnema objektide kontuurid ja siis erinevad ka nende detailid, kuna fotoretseptorite ja visuaalsete neuronite tundlikkus pimedas suureneb järk-järgult.
Valgustundlikkuse suurenemine pimedas viibimise ajal toimub ebaühtlaselt: esimese 10 minutiga suureneb see kümneid kordi ja seejärel tunni jooksul kümneid tuhandeid kordi. Olulist rolli selles protsessis mängib visuaalsete pigmentide taastamine. Kuna pimedas on tundlikud ainult vardad, on nõrgalt valgustatud objekt nähtav ainult perifeerse nägemisega. Kohanemises mängib olulist rolli lisaks visuaalsetele pigmentidele võrkkesta elementide vaheliste ühenduste vahetamine. Pimedas suureneb ganglionraku vastuvõtuvälja ergutuskeskuse pindala rõnga inhibeerimise nõrgenemise tõttu, mis põhjustab valgustundlikkuse suurenemist. Silma valgustundlikkus sõltub ka ajust tulevatest mõjutustest. Ühe silma valgustamine vähendab valgustamata silma valgustundlikkust. Lisaks mõjutavad valgustundlikkust ka heli-, haistmis- ja maitsesignaalid.
2.12. Nägemise diferentsiaalne tundlikkus
Kui lisavalgustus dI langeb valgustatud pinnale heledusega I, siis Weberi seaduse kohaselt märkab inimene valgustuse erinevust ainult siis, kui dI / I \u003d K, kus K on konstant 0,01–0,015. dI/I väärtust nimetatakse valgustundlikkuse diferentsiaalläveks. Suhe dI/I on erinevatel valgustustasemetel konstantne ja tähendab, et kahe pinna valgustuse erinevuse tajumiseks peab üks neist olema teisest 1 - 1,5% võrra heledam.
2.13. Heleduse kontrastsus
Visuaalsete neuronite vastastikune lateraalne inhibeerimine (vt ptk 3) on üldise või globaalse heleduse kontrastsuse aluseks. Seega tundub heledal taustal lebav hall pabeririba tumedam kui sama riba tumedal taustal. Seda seletatakse asjaoluga, et hele taust ergastab paljusid võrkkesta neuroneid ja nende erutus pärsib riba poolt aktiveeritud rakke. Tugevaim külgmine inhibeerimine toimib tihedalt asetsevate neuronite vahel, luues kohaliku kontrasti efekti. Erineva valgustusega pindade piiril on ilmne heleduse erinevuse suurenemine. Seda efekti nimetatakse ka kontuuride täiustamiseks ehk Machi efektiks: ereda valgusvälja ja tumedama pinna piiril on näha kaks lisajoont (veel heledam joon ereda välja piiril ja väga tume joon tumeda pinna piir).
2.14. Valguse pimestav heledus
Liiga ere valgus põhjustab ebameeldiva pimestava tunde. Pimestava heleduse ülempiir sõltub silma kohanemisest: mida pikem oli pimedas kohanemine, seda madalam valguse heledus põhjustab pimestamist. Kui vaatevälja satuvad väga eredad (pimestavad) objektid, halvendavad need signaalide eristamist olulisel osal võrkkestast (näiteks öisel teel pimestavad juhte vastutulevate autode esituled). Silmade pingega seotud delikaatsete tööde puhul (pikk lugemine, arvutiga töötamine, väikeste osade kokkupanek) tuleks kasutada ainult hajutatud valgust, mis ei pimesta silmi.
2.15. Nägemise inerts, värelevate, järjestikuste kujutiste sulandumine
Visuaalne tunne ei ilmu koheselt. Enne aistingu tekkimist peab visuaalses süsteemis toimuma mitu transformatsiooni ja signaalimist. Nägemise inertsi aeg, mis on vajalik nägemisaistingu ilmnemiseks, on keskmiselt 0,03–0,1 s. Tuleb märkida, et ka see tunne ei kao kohe pärast ärrituse lakkamist - see püsib mõnda aega. Kui liigutame pimedas põlevat tikku läbi õhu, näeme helendavat joont, kuna üksteise järel kiiresti järgnevad valgusstiimulid sulanduvad pidevaks aistinguks. Valgusstiimulite (näiteks valgussähvatuste) minimaalset kordumissagedust, mille juures tekib üksikute aistingute seos, nimetatakse kriitiline väreluse sulamissagedus. Keskmise valgustuse korral on see sagedus 10-15 välku 1 sekundis. Kino ja televisioon põhinevad sellel nägemise omadusel: me ei näe üksikute kaadrite vahel lünki (kinos 24 kaadrit 1 s kohta), kuna visuaalne tunnetus ühest kaadrist kestab ikka kuni järgmise ilmumiseni. See loob illusiooni pildi ja selle liikumise järjepidevusest.
Aistingud, mis jätkuvad pärast stimulatsiooni lõppemist, nimetatakse järjestikused pildid. Kui vaatate kaasasolevat lampi ja sulgete silmad, on see mõnda aega nähtav. Kui peale pilgu fikseerimist valgustatud objektile nihutatakse pilk heledale taustale, siis mõnda aega võib sellest objektist näha negatiivset pilti, s.t. selle heledad osad on tumedad ja tumedad heledad (negatiivne järjestikune pilt). Seda seletatakse asjaoluga, et valgustatud objektist tulenev ergastus aeglustab (kohandab) lokaalselt võrkkesta teatud piirkondi; kui pärast seda nihutate pilgu ühtlaselt valgustatud ekraanile, siis selle valgus erutab rohkem neid piirkondi, mis varem ei erutunud.
2.16. värvinägemine
Kogu elektromagnetilise kiirguse spekter, mida me näeme, jääb lühikese lainepikkusega (lainepikkusega 400 nm) kiirguse, mida me nimetame violetseks, ja pikalainelise kiirguse (lainepikkus 700 nm), mida nimetatakse punaseks, vahele. Ülejäänud nähtava spektri värvid (sinine, roheline, kollane ja oranž) on vahepealsete lainepikkustega. Kõikide värvide kiirte segamine annab valge. Seda saab ka kahe nn paaris komplementaarse värvi segamisel: punane ja sinine, kollane ja sinine. Kui segate kolm põhivärvi (punane, roheline ja sinine), saate mis tahes värvi.
Maksimaalset äratundmist naudib G. Helmholtzi kolmekomponendiline teooria, mille kohaselt tagavad värvitaju kolme tüüpi erineva värvitundlikkusega koonused. Mõned neist on tundlikud punase, teised rohelise ja kolmandad sinise suhtes. Iga värv mõjutab kõiki kolme värvitundlikku elementi, kuid erineval määral. See teooria leidis otsest kinnitust katsetes, mille käigus mõõdeti inimese võrkkesta üksikutes koonustes erineva lainepikkusega kiirguse neeldumist.
Osalist värvipimedust kirjeldati 18. sajandi lõpus. D. Dalton, kes ise selle all kannatas. Seetõttu tähistati värvitaju anomaaliat terminiga "värvipimedus". Värvipimedust esineb 8% meestest; seda seostatakse teatud geenide puudumisega meeste sugu määravas paaritu X-kromosoomis. Professionaalses valikus olulise värvipimeduse diagnoosimiseks kasutatakse polükromaatilisi tabeleid. Selle all kannatavad inimesed ei saa olla täisväärtuslikud transpordijuhid, kuna nad ei pruugi eristada fooride ja liiklusmärkide värvi. Osalist värvipimedust on kolme tüüpi: protanoopia, deuteranoopia ja tritanopia. Igaüht neist iseloomustab kolmest põhivärvist ühe taju puudumine. Protanoopia all kannatavad inimesed ("punapimedad") ei taju punast, sini-sinised kiired tunduvad neile värvitud. Deuteranoopiat ("roheline-pimedad") põdevad inimesed ei erista rohelist tumepunasest ja sinisest. Tritanopiaga (haruldane värvinägemise anomaalia) siniseid ja violetseid kiiri ei tajuta. Kõik loetletud osalise värvipimeduse tüübid on kolmekomponendilise teooriaga hästi seletatavad. Igaüks neist on tingitud ühe kolmest koonuse värvi retseptorist puudumisest.
2.17. Ruumi tajumine
nägemisteravus nimetatakse maksimaalseks võimeks eristada objektide üksikuid detaile. Selle määrab kahe punkti vaheline väikseim kaugus, mida silm eristab, s.t. näeb eraldi, mitte koos. Normaalne silm eristab kahte punkti, mille vaheline kaugus on 1 kaareminut. Võrkkesta keskel on maksimaalne nägemisteravus - kollane laik. Selle äärealadel on nägemisteravus palju väiksem. Nägemisteravust mõõdetakse spetsiaalsete tabelite abil, mis koosnevad mitmest tähereast või erineva suurusega avatud ringidest. Tabeli järgi määratud nägemisteravust väljendatakse suhtelistena ja normaalset nägemisteravust võetakse üheks. On inimesi, kellel on üliäge nägemine (visus rohkem kui 2).
Vaateväli. Kui vaatate väikest eset, projitseeritakse selle kujutis võrkkesta kollasele kohale. Sel juhul näeme objekti keskse nägemisega. Selle nurga suurus inimestel on vaid 1,5-2 nurgakraadi. Objekte, mille kujutised langevad ülejäänud võrkkestale, tajub perifeerne nägemine. Nimetatakse ruumi, mis on silmaga nähtav pilku ühte punkti fikseerides vaateväli. Vaatevälja piiri mõõtmine toimub piki perimeetrit. Värvitu objektide vaatevälja piirid on allapoole 70, üles - 60, sissepoole - 60 ja väljapoole - 90 kraadi. Mõlema silma vaateväljad inimestel langevad osaliselt kokku, mis on ruumi sügavuse tajumisel väga oluline. Erinevate värvide vaateväljad ei ole samad ja on väiksemad kui mustvalgete objektide puhul.
binokulaarne nägemine See on nägemine kahe silmaga. Tavalise nägemisega inimesel ei ole mistahes objekti vaadates kahe objekti tunnet, kuigi kahel võrkkestal on kaks kujutist. Selle objekti iga punkti kujutis langeb kahe võrkkesta nn vastavatele ehk vastavatele lõikudele ja inimese tajumisel sulanduvad kaks kujutist üheks. Kui ühele silmale küljelt kergelt vajutada, hakkab see silmades kahekordistuma, sest võrkkesta kirjavahetus on häiritud. Kui vaadata lähedast objekti, siis mõne kaugema punkti kujutis langeb kahe võrkkesta mitteidentsetele (erinevatele) punktidele. Erinevus mängib suurt rolli kauguse hindamisel ja seega ka ruumi sügavuse nägemisel. Inimene suudab märgata sügavuse muutust, mis tekitab võrkkestal pildis mitme kaaresekundi pikkuse nihke. Binokulaarne sulandumine või kahe võrkkesta signaalide ühendamine üheks neuraalseks kujutiseks toimub aju esmases visuaalses ajukoores.
Objekti suuruse hinnang. Tuntud objekti suurust hinnatakse selle võrkkesta kujutise suuruse ja objekti kauguse silmadest funktsioonina. Juhul, kui kaugust võõra objektini on raske hinnata, on selle suuruse määramisel võimalik suuri vigu.
Kauguse hinnang. Ruumi sügavuse tajumine ja kauguse hindamine objektini on võimalik nii ühe silmaga (monokulaarne nägemine) kui ka kahe silmaga (binokulaarne nägemine) nähes. Teisel juhul on kauguse hinnang palju täpsem. Akommodatsiooni fenomen on monokulaarse nägemise korral lähikauguste hindamisel teatud tähtsusega. Kauguse hindamisel on oluline ka see, et tuttava objekti kujutis võrkkestal oleks seda suurem, mida lähemal see on.
Silmade liikumise roll nägemises. Mis tahes objekti vaadates silmad liiguvad. Silmaliigutusi teostavad 6 silmamuna külge kinnitatud lihast. Kahe silma liikumine toimub samaaegselt ja sõbralikult. Lähedasi objekte silmas pidades on vaja vähendada (lähenemine) ja kaugemate objektide puhul - eraldada kahe silma visuaalsed teljed (lahknemine). Silmade liigutuste olulise rolli nägemisele määrab ka see, et aju visuaalse info pidevaks vastuvõtmiseks on vaja pilti võrkkestal liigutada. Impulsid nägemisnärvis tekivad valguspildi sisse- ja väljalülitamise hetkel. Valguse jätkuval toimel samadele fotoretseptoritele lakkavad impulsid nägemisnärvi kiududes kiiresti ning visuaalne tunne liikumatute silmade ja objektidega kaob 1–2 sekundi pärast. Kui silmale asetada pisikese valgusallikaga iminapp, näeb inimene seda alles siis, kui see sisse või välja lülitatakse, kuna see stiimul liigub koos silmaga ja on seetõttu võrkkesta suhtes liikumatu. Sellise kohanemise (kohanemise) liikumatu pildiga ületamiseks tekitab silm mis tahes objekti vaatamisel pidevaid hüppeid (sakkaade), mis on inimesele märkamatud. Iga hüppe tulemusena nihkub võrkkesta kujutis ühelt fotoretseptorilt teisele, põhjustades taas ganglionrakkude impulsse. Iga hüppe kestus on võrdne sajandiksekundiga ja selle amplituud ei ületa 20 nurkkraadi. Mida keerulisem on vaatlusalune objekt, seda keerulisem on silmade liikumise trajektoor. Need justkui "jälgivad" pildi kontuure (joonis 4.6), jäädes selle kõige informatiivsematele aladele (näiteks näos, need on silmad). Lisaks hüpetele värisevad ja triivivad silmad pidevalt peenelt (nihkuvad aeglaselt pilgu fikseerimise kohast). Need liigutused on väga olulised ka visuaalse taju jaoks.

Riis. 4.6. Silma liikumise trajektoor (B) Nefertiti kujutise uurimisel (A)
Inimeste ja paljude loomade võrkkesta vardad sisaldavad pigmenti rodopsiini ehk visuaallillat, mille koostist, omadusi ja keemilisi muundumisi on viimastel aastakümnetel põhjalikult uuritud. Koonustest leiti pigment jodopsiini. Koonused sisaldavad ka pigmente chlorolab ja erythrolab; esimene neist neelab kiired, mis vastavad rohelisele, ja teine - spektri punane osa.
Rodopsiin on suure molekulmassiga ühend (molekulmass 270 000), mis koosneb võrkkesta - A-vitamiini aldehüüdist ja opsiinkiirest. Valguskvanti toimel toimub selle aine fotofüüsikaliste ja fotokeemiliste transformatsioonide tsükkel: võrkkesta isomeriseerub, selle kõrvalahel sirgub, side võrkkesta ja valgu vahel katkeb ning valgumolekuli ensümaatilised keskused aktiveeruvad. Konformatsiooniline muutus pigmendi molekulides aktiveerib Ca2+ ioonid, mis jõuavad difusiooni teel naatriumikanalitesse, mille tulemusena väheneb juhtivus Na+ suhtes. Naatriumi juhtivuse vähenemise tulemusena toimub fotoretseptori raku sees rakuvälise ruumi suhtes elektronegatiivsuse suurenemine. Seejärel eraldatakse võrkkesta opsiinist. Reetina reduktaasi nimelise ensüümi mõjul muutub viimane A-vitamiiniks.
Silmade tumenemisel toimub visuaalse lilla taastumine, s.t. rodopsiini resüntees. See protsess nõuab, et võrkkest saaks A-vitamiini cis-isomeeri, millest moodustub võrkkesta. Kui organismis puudub A-vitamiin, on rodopsiini moodustumine järsult häiritud, mis viib ööpimeduse tekkeni.
Fotokeemilised protsessid võrkkestas toimuvad väga vähe; isegi väga ereda valguse toimel lõheneb vaid väike osa pulgas leiduvast rodopsiinist.
Jodopsiini struktuur on lähedane rodopsiini omale. Jodopsiin on ka võrkkesta ühend valgu opsiiniga, mida toodetakse koonustes ja mis erineb varraste opsiinist.
Valguse neeldumine rodopsiini ja jodopsiini poolt on erinev. Jodopsiin neelab kõige suuremal määral kollast valgust lainepikkusega umbes 560 nm.
Võrkkesta on üsna keeruline närvivõrk, millel on horisontaalsed ja vertikaalsed ühendused fotoretseptorite ja rakkude vahel. Bipolaarsed võrkkesta rakud edastavad signaale fotoretseptoritelt ganglionrakkude kihti ja amakriinrakkudesse (vertikaalne ühendus). Horisontaalsed ja amakriinsed rakud osalevad horisontaalses signaaliülekandes naaberfotoretseptorite ja ganglionrakkude vahel.
Võrkkesta elektrilised nähtused äratasid teadlaste tähelepanu pärast võrkkesta potentsiaalsete erinevuste kõikumiste avastamist sõltuvalt selle valgustuse tingimustest. Selle protsessi salvestamist nimetatakse elektroretinogrammiks (ERG). Võrkkesta valgustundlike elementide uurimise oluline meetod on nägemisnärvi üksikute kiudude elektrilise aktiivsuse registreerimine silma valguse mõjul. See meetod võimaldas tuvastada kolme peamise valgustundlike elementide rühma olemasolu. Esimene neist saadab impulsse kogu valgusstiimuli toimeaja jooksul, paljastades valgusega kohanemisel nende sageduse vähese vähenemise. Teine on põnevil ja saadab seetõttu impulsse ainult siis, kui silm on valgustatud ja tumenenud. Kolmas rühm reageerib erutusega ainult elektrikatkestusele; selle kategooria valgustundlikud elemendid saadavad impulsse pimeduse ajal ja neid pärsib silma valgustus. Kõiki kolmest loetletud võrkkesta fotoretseptorite rühmast iseloomustab sellele rühmale iseloomulik elektrilise oleku muutus, kui silm on valgustatud; ERG on võrkkesta kõigist kolmest elektrilisest protsessist tulenev summakõver. Võrkkesta varraste elementide hulgas domineerivad I rühma valgustundlikud elemendid. Koonused on peamiselt II ja III rühma fotoretseptorid. Võrkkestas vabanev elektrienergia tuleneb selles toimuvatest ainevahetusprotsessidest.
Visuaalse süsteemi keskuste elektriline aktiivsus. Elektrilised nähtused võrkkestas ja nägemisnärvis. Valguse toimel retseptorites ja seejärel võrkkesta neuronites tekivad elektrilised potentsiaalid, mis peegeldavad toimiva stiimuli parameetreid.
Võrkkesta kogu elektrilist reaktsiooni valgusele nimetatakse elektroretinogrammiks (ERG). Seda saab salvestada kogu silmast või otse võrkkestast. Selleks asetatakse üks elektrood sarvkesta pinnale ja teine - näonahale silma lähedal või kõrvapulgale. Elektroretinogrammil eristatakse mitmeid iseloomulikke laineid (joon. 14.8). Laine a peegeldab fotoretseptorite sisemiste segmentide (hilise retseptori potentsiaal) ja horisontaalsete rakkude ergastust. Laine b tekib võrkkesta gliaalrakkude (Mülleri) aktiveerimise tulemusena bipolaarsete ja amakriinsete neuronite ergastamisel vabanevate kaaliumiioonide poolt. Laine c peegeldab pigmendi epiteelirakkude aktiveerumist ja laine d peegeldab horisontaalsete rakkude aktiveerumist.
Valgusstiimuli intensiivsus, värvus, suurus ja kestus kajastuvad ERG-s hästi. Kõigi ERG lainete amplituud suureneb võrdeliselt valguse intensiivsuse logaritmiga ja ajaga, mille jooksul silm oli pimedas. Laine d (reaktsioon väljalülitamisele) on seda suurem, mida kauem tuli põles. Kuna ERG peegeldab peaaegu kõigi võrkkesta rakkude aktiivsust (v.a ganglionrakud), kasutatakse seda indikaatorit laialdaselt silmahaiguste kliinikus erinevate võrkkesta haiguste diagnoosimiseks ja ravi kontrollimiseks.
Võrkkesta ganglionrakkude ergastamine toob kaasa asjaolu, et impulsid sööstavad mööda nende aksoneid (nägemisnärvi kiud) ajju. Võrkkesta ganglionrakk on esimene "klassikalist" tüüpi neuron fotoretseptori-aju ahelas. Kirjeldatud on kolme peamist ganglionrakkude tüüpi: reageerivad sisselülitamisele (sissereaktsioon), valguse väljalülitamisele (välja-reaktsioon) ja mõlemad (sisse-välja-reaktsioon) (joon. 14.9).
Võrkkesta keskosas olevate ganglionrakkude retseptsiooniväljade läbimõõt on palju väiksem kui perifeerias. Need vastuvõtlikud väljad on ümmargused ja kontsentriliselt ehitatud: ümmargune ergastav keskpunkt ja rõngakujuline pärssivat perifeerne tsoon või vastupidi. Retseptiivse välja keskel vilkuva valgustäpi suuruse suurenemisega suureneb ganglionraku reaktsioon (ruumiline summeerimine). Lähedal asuvate ganglionrakkude samaaegne ergastamine põhjustab nende vastastikust inhibeerimist: iga raku vastused muutuvad väiksemaks kui ühe stimulatsiooni korral. See toime põhineb lateraalsel või lateraalsel inhibeerimisel. Naaberganglionrakkude vastuvõtlikud väljad kattuvad osaliselt, nii et samad retseptorid võivad osaleda vastuste genereerimises mitmelt neuronilt. Võrkkesta ganglionrakkude retseptiivsed väljad toodavad oma ümara kuju tõttu võrkkesta kujutise nn punkt-punkti kirjelduse: seda kuvab väga õhuke ergastatud neuronitest koosnev mosaiik.
Võrkkestast eemaldatud elektrilist kogupotentsiaali nimetatakse elektroretinogrammiks. Seda saab registreerida, "kandes ühe elektroodi sarvkesta pinnale ja teise silma lähedal olevale nahale. See potentsiaal peegeldab pigmendirakkude ja fotoretseptorite plasmamembraani läbivate elektrivoolude summat. Arvatakse, et a-laine on retseptori potentsiaalide summa, b-laine peegeldab gliiarakkude membraanipotentsiaalide muutust, e-laine - pigmendi epiteelirakud, d-laine tekib võrkkesta neuronite membraanipotentsiaalide muutumise tõttu.
Silma sisemine kest - võrkkest - on visuaalse analüsaatori retseptori osa, milles toimub valguse tajumine ja visuaalsete aistingute esmane analüüs. Sarvkesta, läätse, klaaskeha ja kogu võrkkesta paksuse läbiv valguskiir tabab esmalt välimist (pupillikihist kõige kaugemal asuvat pigmendiepiteelirakkudest. Nendes rakkudes paiknev pigment neelab valgust, takistades seeläbi selle peegeldus ja dispersioon, mis aitab kaasa taju selgusele Fotoretseptori rakud-pulgad ja koonused külgnevad seestpoolt pigmendikihiga, paiknevad ebaühtlaselt (kollase laigu piirkonnas paiknevad ainult koonused, vähendavad koonused perifeeria suunas, ja suurendavad varraste arvu) Vardad vastutavad hämaras nägemise eest, koonused värvuse eest .Mikroskoobiliselt on võrkkest 3 neuronist koosnev ahel: fotoretseptorid-välimine neuron, assotsiatiivne-keskmine, ganglion-sisemine. närviimpulss 1-2 neuronilt saadakse välise (pleksikujulise) kihi sünapside kaudu.rakk ja teine ganglionrakkude dentriididega Bipolaarsed rakud on kontaktis mitme pulgaga ja ainult ühe koonusega Ühe rakuga ühendatud fotoretseptorid moodustavad retseptiivse Kolmandate rakkude aksonid, kui need on ühendatud, moodustavad nägemisnärvi tüve.
Fotokeemilised protsessid võrkkestas. Võrkkesta retseptorrakud sisaldavad valgustundlikke pigmente – kompleksseid valkaineid, kromoproteiine, mis valguse käes värvuvad. Välimiste segmentide membraanil olevad vardad sisaldavad rodopsiini, koonused sisaldavad jodopsiini ja muid pigmente. Rodopsiin ja jodopsiin koosnevad võrkkesta (A-vitamiini aldehüüd) ja opsiini glükoproteiinist.
Kui A-vitamiini sisaldus organismis väheneb, nõrgenevad rodopsiini resünteesi protsessid, mis põhjustab hämaras nägemise halvenemist - nn "ööpimedust". Pideva ja ühtlase valgustusega saavutatakse tasakaal pigmentide lagunemiskiiruse ja taassünteesi vahel. Kui võrkkestale langeva valguse hulk väheneb, on see dünaamiline tasakaal häiritud ja nihkub kõrgemate pigmendikontsentratsioonide suunas. See fotokeemiline nähtus on pimedas kohanemise aluseks.
Fotokeemilistes protsessides on eriti oluline võrkkesta pigmendikiht, mille moodustab fustsiini sisaldav epiteel. See pigment neelab valgust, takistades selle peegeldumist ja hajumist, mis tagab visuaalse taju selguse. Pigmendirakkude protsessid ümbritsevad varraste ja koonuste valgustundlikke segmente, osaledes fotoretseptorite metabolismis ja visuaalsete pigmentide sünteesis.
Silma fotoretseptorites tekib fotokeemiliste protsesside tõttu valguse toimel retseptori potentsiaal retseptori membraani hüperpolarisatsiooni tõttu. See on visuaalsete retseptorite eripära, teiste retseptorite aktiveerimine väljendub nende membraani depolarisatsioonis. Visuaalse retseptori potentsiaali amplituud suureneb koos valgusstiimuli intensiivsuse suurenemisega.
silmade liigutused mängivad visuaalses tajumises väga olulist rolli. Isegi juhul, kui vaatleja fikseerib oma pilguga kindla punkti, ei ole silm paigal, vaid teeb kogu aeg väikseid liigutusi, mis on tahtmatud. Silmade liigutused täidavad statsionaarsete objektide vaatamisel kohanematuse funktsiooni. Väikeste silmade liigutuste teine funktsioon on hoida kujutist selge nägemise tsoonis.
Nägemissüsteemi reaalsetes tingimustes liiguvad silmad kogu aeg, uurides nägemisvälja kõige informatiivsemaid osi. Samal ajal võimaldavad mõned silmaliigutused arvestada vaatlejast samal kaugusel asuvate objektidega, näiteks pilti lugedes või vaadates, teised - temast erineval kaugusel asuvate objektide vaatamisel. Esimest tüüpi liigutused on mõlema silma ühesuunalised liigutused, teist tüüpi aga koondavad või eraldavad nägemisteljed, s.t. liikumised on suunatud vastassuundades.
On näidatud, et silmade ülekandumise ühelt objektilt teisele määrab nende teabesisu. Pilk ei peatu nendel aladel, mis sisaldavad vähe infot ja samas fikseerib pikaks ajaks kõige informatiivsemad kohad (näiteks objekti kontuurid). See funktsioon on kahjustatud, kui otsmikusagarad on kahjustatud. Silmade liikumine võimaldab tajuda objektide üksikuid tunnuseid, nende vahekorda, mille põhjal moodustub tervikpilt, mis salvestatakse pikaajalisse mällu.
14.1.6. Sensoorsete süsteemide koostoime
Sensoorsete süsteemide koostoime toimub seljaaju, retikulaarse, talamuse ja kortikaalsel tasemel. Signaalide integratsioon retikulaarses formatsioonis on eriti lai. Ajukoores toimub kõrgema järgu signaalide integratsioon. Mitmete ühenduste moodustumise tulemusena teiste sensoorsete ja mittespetsiifiliste süsteemidega omandavad paljud kortikaalsed neuronid võime reageerida erineva modaalsusega signaalide keerukatele kombinatsioonidele. See kehtib eriti assotsiatsioonipiirkondade närvirakkude kohta. koor poolkerad, millel on kõrge plastilisus, mis tagab nende ümberkorraldamise
omadused pideva õppimise protsessis uute stiimulite äratundmiseks. Intersensoorne (ristmodaalne) interaktsioon kortikaalsel tasandil loob tingimused "maailma skeemi (või kaardi)" kujunemiseks ja keha enda "kehaskeemi" pidevaks sidumiseks, koordineerimiseks sellega.
14.2. SENSORSÜSTEEMIDE ERIFÜSIOLOOGIA
14.2.1. visuaalne süsteem
Nägemine on evolutsiooniliselt kohanenud elektromagnetilise kiirguse tajumiseks nende leviala teatud, väga kitsas osas (nähtav valgus). Visuaalne süsteem annab ajule üle 90% sensoorsest teabest. Nägemine on mitmelüliline protsess, mis algab kujutise projitseerimisega ainulaadse perifeerse optilise seadme – silma – võrkkestale. Seejärel toimub fotoretseptorite ergastamine, visuaalse teabe edastamine ja muundamine visuaalse süsteemi närvikihtides ning visuaalne tajumine lõpeb visuaalse kujutise otsuse vastuvõtmisega selle süsteemi kõrgemate kortikaalsete osade poolt.
Silma optilise aparaadi ehitus ja funktsioonid. Silmal on sfääriline kuju, mis hõlbustab kõnealust objekti sihtimiseks pööramist. Teel silma valgustundlikule membraanile (võrkkestani) läbivad valguskiired mitut läbipaistvat keskkonda - sarvkesta, läätse ja klaaskeha. Silmasiseste valguskiirte murdumise määrab sarvkesta ja vähesel määral ka läätse teatud kumerus ja murdumisnäitaja (joon. 14.2).
Iga optilise süsteemi murdumisvõimet väljendatakse dioptrites (D). Üks diopter võrdub 100 cm fookuskaugusega läätse murdumisvõimega.Terve silma murdumisvõime on kaugel asuvate objektide vaatamisel 59D ja lähedal asuvate objektide vaatamisel 70,5D. Objekti kujutise võrkkestale projitseerimise skemaatiliseks esitamiseks peate tõmbama selle otstest läbi sõlmepunkti (7 mm sarvisest tagapool) jooned.
kestad). Võrkkestale saadakse pilt, mis on järsult vähendatud ja pööratud tagurpidi ja paremalt vasakule (joon. 14.3).
Majutus. Akommodatsiooni nimetatakse silma kohanemiseks erinevatel vahemaadel asuvate objektide selge nägemisega. Objekti selgeks nägemiseks on vajalik, et see oleks fokusseeritud võrkkestale, see tähendab, et selle pinna kõikidest punktidest saadavad kiired projitseeritakse võrkkesta pinnale (joonis 14.4). Kui vaatame kaugeid objekte (A), on nende kujutis (a) fokuseeritud võrkkestale ja need on selgelt nähtavad. Kuid lähedalasuvate objektide (B) pilt (b) on udune, kuna nende kiired kogutakse võrkkesta taha. Akommodatsioonis mängib põhirolli lääts, mis muudab selle kumerust ja sellest tulenevalt ka murdumisvõimet. Lähedasi objekte vaadates muutub lääts kumeramaks (vt joon. 14.2), mille tõttu objekti mis tahes punktist lahknevad kiired koonduvad võrkkestale. Akommodatsioonimehhanismiks on tsiliaarsete lihaste kokkutõmbumine, mis muudab läätse kumerust. Lääts on ümbritsetud õhukese läbipaistva kapsliga, mis on alati venitatud, st lamestatud tsiliaarse vöö (tsinn ligament) kiudude abil. Siliaarkeha silelihasrakkude kokkutõmbumine vähendab tsooni sidemete tõmmet, mis suurendab läätse kumerust selle elastsuse tõttu. Tsiliaarseid lihaseid innerveerivad okulomotoorse närvi parasümpaatilised kiud. Atropiini sattumine silma põhjustab erutuse ülekandumise sellesse lihasesse, piirab silma majutust lähedaste objektide vaatamisel. Vastupidi, parasümpatomimeetilised ained - pilokarpiin ja ezeriin - põhjustavad selle lihase kokkutõmbumist.
Noore inimese normaalse silma jaoks asub selge nägemise kaugeim punkt lõpmatuseni. Ta uurib kaugeid objekte ilma akommodatsioonipingeta, st ilma kokkutõmbumiseta.
tsiliaarne lihas. Lähim selge nägemise punkt on silmast 10 cm kaugusel.
Presbüoopia. Lääts kaotab vanusega oma elastsuse ja kui tsinni sidemete pinge muutub, muutub selle kõverus vähe. Seetõttu ei asu nüüd lähim selge nägemispunkt silmast 10 cm kaugusel, vaid eemaldub sellest. Lähedased objektid ei ole samal ajal nähtavad. Seda seisundit nimetatakse seniilseks kaugnägelikkuseks või presbüoopia. Eakad inimesed on sunnitud kasutama kaksikkumerate läätsedega prille.
Refraktsioonianomaaliad silmad. Silma murdumise kaks peamist anomaaliat – lühinägelikkus ehk lühinägelikkus ja kaugnägelikkus ehk hüpermetroopia – ei tulene mitte silma murdumiskeskkonna puudulikkusest, vaid silmamuna pikkuse muutumisest (joon. 14.5. A).
Lühinägelikkus. Kui silma pikitelg on liiga pikk, siis kaugemal asuva objekti kiired ei keskendu mitte võrkkestale, vaid selle ette, klaaskehasse (joon. 14.5, B). Sellist silma nimetatakse lühinägelikuks või lühinägelikuks. Selgeks kaugusesse nägemiseks on vaja lühinägelike silmade ette asetada nõgusad prillid, mis viivad fokuseeritud kujutise võrkkestale (joon. 14.5, B).
Kaugnägelikkus. Lühinägelikkuse vastand on kaugnägelikkus ehk hüpermetroopia. Kaugnägevas silmas (joon. 14.5, D) on silma pikitelg lühenenud ja seetõttu on kauge objekti kiired fokusseeritud mitte võrkkestale, vaid selle taha. Seda murdumise puudumist saab kompenseerida kohandatava pingutusega, st läätse kumeruse suurendamisega. Seetõttu pingutab kaugelenägev inimene akommodatiivset lihast, pidades silmas mitte ainult lähedasi, vaid ka kaugeid objekte. Lähedasi objekte silmas pidades on kaugnägeliku kohanemisvõimeline pingutus
neist ei piisa. Seetõttu peaksid kaugnägelikud inimesed lugemiseks kandma kaksikkumerate läätsedega prille, mis suurendavad valguse murdumist (joon. 14.5, E). Hüpermetroopiat ei tohi segi ajada seniilse kaugnägelikkusega. Ainus ühine joon on see, et on vaja kasutada kaksikkumerate läätsedega prille.
Astigmatism. Astigmatism ehk kiirte ebavõrdne murdumine eri suundades (näiteks piki horisontaalset ja vertikaalset meridiaani) kuulub samuti murdumisvigade hulka. Astigmatism ei ole tingitud sarvkesta rangelt sfäärilisest pinnast. Tugeva astme astigmatismi korral võib see pind läheneda silindrilisele, mida korrigeerivad silindrilised prillid, mis kompenseerivad sarvkesta puudusi.
Pupill ja pupillide refleks. Pupill on iirise keskel asuv auk, mille kaudu valguskiired silma sisenevad. Pupill teravdab võrkkesta kujutist, suurendades silma teravussügavust. Läbides ainult keskkiiri, parandab see võrkkesta pilti, kõrvaldades ka sfäärilise aberratsiooni. Kui katate silma valguse eest ja seejärel avate selle, siis tumenemise ajal laienenud pupill kitseneb kiiresti ("pupillirefleks"). Iirise lihased muudavad pupilli suurust, reguleerides silma siseneva valguse hulka. Nii et väga eredas valguses on pupilli minimaalne läbimõõt (1,8 mm), keskmises päevavalguses see laieneb (2,4 mm) ja pimedas maksimaalne (7,5 mm). See toob kaasa võrkkesta kujutise kvaliteedi halvenemise, kuid suurendab nägemise tundlikkust. Pupilli läbimõõdu piirav muutus muudab selle pindala umbes 17 korda. Samal ajal muutub valgusvoog samas ulatuses. Valgustuse intensiivsuse ja pupilli läbimõõdu vahel on logaritmiline seos. Õpilase reaktsioonil valgustuse muutustele on adaptiivne iseloom, kuna see stabiliseerib võrkkesta valgustust väikeses vahemikus.
Iirises on pupilli ümbritsevad kahte tüüpi lihaskiud: ümmargused (m. sphincter iridis), mida innerveerivad okulomotoorse närvi parasümpaatilised kiud, ja radiaalsed (m. dilatator iridis), mida innerveerivad sümpaatilised närvid. Esimese kokkutõmbumine põhjustab ahenemist, teise kokkutõmbumine - õpilase laienemist. Vastavalt sellele põhjustavad atsetüülkoliin ja ezeriin õpilase ahenemist ja adrenaliin - õpilase laienemist. Pupillid laienevad valu, hüpoksia, aga ka sümpaatilise süsteemi erutust suurendavate emotsioonide (hirm, raev) ajal. Pupillide laienemine on mitmete patoloogiliste seisundite, nagu valušokk, hüpoksia, oluline sümptom.
Tervetel inimestel on mõlema silma pupillide suurus sama. Kui üks silm on valgustatud, kitseneb ka teise silmapupill; sellist reaktsiooni nimetatakse sõbralikuks. Mõnel patoloogilisel juhul on mõlema silma pupillide suurus erinev (anisokoria).
Võrkkesta ehitus ja funktsioonid. Võrkkesta on silma sisemine valgustundlik membraan. Sellel on keeruline mitmekihiline struktuur (joonis 14.6). Sekundaarseid fotoretseptoreid on kahte tüüpi, mis erinevad oma funktsionaalse tähtsuse poolest (varras ja koonus) ja mitut tüüpi närvirakke. Fotoretseptorite ergastamine aktiveerib võrkkesta esimese närviraku (bipolaarne neuron). Bipolaarsete neuronite ergastamine aktiveerib võrkkesta ganglionrakud, mis edastavad oma impulsssignaalid subkortikaalsetesse nägemiskeskustesse. Horisontaalsed ja amakriinsed rakud osalevad ka võrkkesta teabe edastamise ja töötlemise protsessides. Moodustuvad kõik ülaltoodud võrkkesta neuronid koos nende protsessidega silma närvisüsteem mis mitte ainult ei edasta informatsiooni aju nägemiskeskustesse, vaid osaleb ka selle analüüsis ja töötlemises. Seetõttu nimetatakse võrkkesta perifeeriasse paigutatud aju osaks.
Nägemisnärvi väljumispunkt silmamunast on nägemisnärvi ketas, mida nimetatakse pimealaks. See ei sisalda fotoretseptoreid ja on seetõttu valguse suhtes tundlik. Me ei tunne võrkkestas "augu" olemasolu.
Vaatleme võrkkesta kihtide struktuuri ja funktsioone, lähtudes võrkkesta välimisest (tagumisest, pupillist kõige kaugemal asuvast) kihist selle kihi sisemise (pupillile lähemal asuva) kihini.
pigmendikiht. Selle kihi moodustab üks rida epiteelirakke, mis sisaldavad suurt hulka erinevaid rakusiseseid organelle, sealhulgas melanosoome, mis annavad sellele kihile musta värvi. See pigment, mida nimetatakse ka varjestuspigmendiks, neelab selleni jõudva valguse, takistades seeläbi selle peegeldumist ja hajumist, mis aitab kaasa visuaalse taju selgusele. Pigmentepiteeli rakkudes on palju protsesse, mis ümbritsevad tihedalt varraste ja koonuste valgustundlikke välissegmente. Pigmendiepiteel mängib otsustavat rolli mitmes funktsioonis, sealhulgas visuaalse pigmendi taassüntees (regenereerimine) pärast selle värvimuutust, fagotsütoosi ja seedimise korral. varraste ja koonuste välimiste segmentide fragmendid.koonused ehk teisisõnu nägemisrakkude välimiste segmentide pideva uuendamise mehhanismis, nägemisrakkude kaitsmisel valguskahjustuste ohu eest, samuti hapniku ja muud ained, mida nad fotoretseptoritele vajavad. Tuleb märkida, et pigmendi epiteeli rakkude ja fotoretseptorite vaheline kontakt on üsna nõrk. Just selles kohas toimub võrkkesta irdumine - ohtlik silmahaigus. Võrkkesta irdumine põhjustab nägemiskahjustusi mitte ainult selle nihkumise tõttu pildi optilise teravustamise kohast, vaid ka retseptorite degeneratsiooni tõttu pigmendiepiteeliga kokkupuute tõttu, mis põhjustab retseptorite metabolismi tõsiseid häireid. ise. Ainevahetushäireid süvendab asjaolu, et toitainete kohaletoimetamine kapillaaridest on häiritud.
silma soonkesta ja fotoretseptorite kiht ise ei sisalda kapillaare (avaskulariseerunud).
Fotoretseptorid. Pigmendikihiga külgneb seestpoolt fotoretseptorite kiht: vardad ja koonused. Iga inimese silma võrkkestas on 6-7 miljonit koonust ja 110-123 miljonit varrast. Need on võrkkestas jaotunud ebaühtlaselt. Võrkkesta keskne fovea (fovea centralis) sisaldab ainult käbisid (kuni 140 tuhat 1 mm 2 kohta). Võrkkesta perifeeria suunas nende arv väheneb ja varraste arv suureneb, nii et kaugemal on ainult vardad. Koonused toimivad kõrge valgustuse tingimustes, tagavad päeva- ja värvinägemise; palju valgustundlikumad vardad vastutavad hämara nägemise eest.
Värvi tajutakse kõige paremini, kui valgus tabab võrkkesta fovea, kus koonused asuvad peaaegu eranditult. Siin on suurim nägemisteravus. Võrkkesta keskpunktist eemaldudes halveneb värvitaju ja ruumiline eraldusvõime järk-järgult. Võrkkesta perifeeria, kus asuvad ainult vardad, ei taju värve. Teisest küljest on võrkkesta koonusaparaadi valgustundlikkus mitu korda väiksem kui vardal, mistõttu hämaras "koonuse" nägemise järsu vähenemise ja "perifeerse" nägemise ülekaalu tõttu saame me hämaras. ei erista värvi ("kõik kassid on öösel hallid").
Pulkade funktsiooni rikkumine, mis tekib A-vitamiini puudusel toidus, põhjustab hämaras nägemise häiret - nn ööpimedust: inimene läheb õhtuhämaruses täiesti pimedaks, päeval aga nägemine püsib. normaalne. Vastupidi, koonuste kahjustamisel tekib fotofoobia: inimene näeb väheses valguses, kuid eredas valguses muutub pimedaks. Sellisel juhul võib tekkida ka täielik värvipimedus – akromaasia.
Fotoretseptori raku struktuur. Fotoretseptori rakk – varras või koonus – koosneb visuaalset pigmenti sisaldavast valgustundlikust välimisest segmendist, sisesegmendist, ühendusjalast, suure tuumaga tuumaosast ja presünaptilisest otsast. Võrkkesta varras ja koonus on nende valgustundlike välimiste segmentide abil pööratud pigmendiepiteeli poole, st valguse vastassuunas. Kell Inimestel sisaldab fotoretseptori välimine segment (pulk või koonus) umbes tuhat fotoretseptori ketast. Varda välimine segment on palju pikem kui koonused ja sisaldab rohkem visuaalset pigmenti. See seletab osaliselt varda suuremat valgustundlikkust: varras
suudab ergastada ainult ühe valguskvanti ja koonuse aktiveerimiseks kulub rohkem kui sada footoni.
Fotoretseptori ketta moodustavad kaks servadest ühendatud membraani. Kettamembraan on tüüpiline bioloogiline membraan, mille moodustab fosfolipiidimolekulide topeltkiht, mille vahel on valgumolekulid. Ketta membraan on rikas polüküllastumata rasvhapete poolest, mis põhjustab selle madala viskoossuse. Selle tulemusena pöörlevad selles olevad valgumolekulid kiiresti ja liiguvad aeglaselt mööda ketast. See võimaldab valkudel sageli põrkuda ja interaktsioonil lühikese aja jooksul moodustada funktsionaalselt olulisi komplekse.
Fotoretseptori sisemine segment on ühendatud välimise segmendiga modifitseeritud tsiliumiga, mis sisaldab üheksa paari mikrotuubuleid. Sisemises segmendis on suur tuum ja kogu raku metaboolne aparaat, sealhulgas mitokondrid, mis tagavad fotoretseptori energiavajaduse, ning valgusünteesi süsteem, mis tagab välissegmendi membraanide uuenemise. Siin toimub visuaalsete pigmendimolekulide süntees ja inkorporeerimine ketta fotoretseptori membraani. Tunni jooksul moodustub sisemise ja välimise segmendi piiril keskmiselt kolm uut ketast. Seejärel liiguvad nad aeglaselt (inimesel umbes 2-3 nädala jooksul) pulga välissegmendi põhjast selle tippu. Lõpuks katkeb välissegmendi tipp, mis sisaldab kuni sada praegu vana ketast ja fagotsütoositakse pigmendikihi rakkude poolt. See on üks olulisemaid mehhanisme fotoretseptori rakkude kaitsmiseks nende valguse eluea jooksul kogunevate molekulaarsete defektide eest.
Ka koonuste välimised segmendid uuenevad pidevalt, kuid aeglasemalt. Huvitaval kombel on olemas ööpäevane uuenemisrütm: varraste välimiste segmentide tipud murduvad peamiselt ära ja fagotsüteeritakse hommikul ja päeval ning koonused - õhtul ja öösel.
Retseptori presünaptiline ots sisaldab sünaptilist linti, mille ümber on palju glutamaati sisaldavaid sünaptilisi vesiikuleid.
visuaalsed pigmendid. Inimese võrkkesta vardad sisaldavad pigmenti rodopsiini ehk visuaalselt lillat, mille maksimaalne neeldumisspekter jääb 500 nanomeetri (nm) vahemikku. Kolme tüüpi koonuste (sinise-, rohelise- ja punase-tundlikud) välimised segmendid sisaldavad kolme tüüpi visuaalseid pigmente, mille neeldumisspektri maksimumid on sinises (420 nm), rohelises (531 nm) ja punases ( 558 nm) spektri osad. Punast koonuse pigmenti nimetatakse jodo-psiiniks. Visuaalse pigmendi molekul on suhteliselt väike (molekulmassiga umbes 40 kilodaltonit), koosneb suuremast valguosast (opsiin) ja väiksemast kromofooriosast (võrkkesta ehk A-vitamiini aldehüüd). Võrkkesta võib olla erinevates
ruumilised konfiguratsioonid, st isomeersed vormid, kuid ainult üks neist, võrkkesta 11-cis-isomeer, toimib kõigi teadaolevate visuaalsete pigmentide kromofoorrühmana. Organismi võrkkesta allikaks on karotenoidid, mistõttu nende puudus põhjustab A-vitamiini vaegust ja selle tulemusena rodopsiini ebapiisavat resünteesi, mis omakorda põhjustab hämaruse nägemise halvenemist ehk “ööpimedust”. Fotoretseptsiooni molekulaarfüsioloogia. Mõelge varda välimise segmendi molekulide muutuste järjestusele, mis vastutab selle ergastamise eest (joonis 14.7, A). Kui valguskvant neelab visuaalse pigmendi molekuli (rodopsiin), isomeriseerub selle kromofoorrühm koheselt: 11-cis-võrkkest sirgub ja muutub täis-trans-võrkkestaks. See reaktsioon kestab umbes 1 ps (1–12 s). Valgus toimib päästikuna või päästikuna, mis käivitab fotovastuvõtu mehhanismi. Võrkkesta fotoisomerisatsioonile järgnevad ruumilised muutused molekuli valguosas: see muutub värvituks ja muundub metarodopsiin II olekusse. Selle tulemusena kinnitub visuaalse pigmendi molekul
omandab võime suhelda teise valguga – membraaniga seotud guanosiintrifosfaati siduva valgu transduktiiniga (T). Kompleksis metarodopsiin II-ga muutub transdutsiin aktiivseks ja vahetab pimedas sellega seotud guanosiindifosfaadi (GDP) guanosiintrifosfaadi (GTP) vastu. Metarodopsiin II on võimeline aktiveerima umbes 500-1000 transdutsiini molekuli, mis viib valgussignaali suurenemiseni.
Iga aktiveeritud transdutsiini molekul, mis on seotud GTP molekuliga, aktiveerib teise membraaniga seotud valgu, fosfodiesteraasi ensüümi (PDE) ühe molekuli. Aktiveeritud PDE hävitab suure kiirusega tsüklilise guanosiinmonofosfaadi (cGMP) molekulid. Iga aktiveeritud PDE molekul hävitab mitu tuhat cGMP molekuli – see on järjekordne samm signaali võimendamisel fotovastuvõtumehhanismis. Kõigi kirjeldatud valguskvanti neeldumisest põhjustatud sündmuste tagajärg on vaba cGMP kontsentratsiooni langus retseptori välimise segmendi tsütoplasmas. See omakorda toob kaasa ioonikanalite sulgumise välissegmendi plasmamembraanis, mis avanesid pimedas ja mille kaudu Na + ja Ca 2+ rakku sisenesid. Ioonikanal sulgub tänu sellele, et vaba cGMP kontsentratsiooni languse tõttu rakus lahkuvad kanalist cGMP molekulid, mis olid sellega pimedas seotud ja hoidsid seda avatuna.
Na + välimisse segmenti sisenemise vähenemine või peatumine põhjustab rakumembraani hüperpolarisatsiooni, st retseptori potentsiaali ilmnemist sellel. Joonisel fig. 14.7, B näitab ioonvoolude suundi, mis pimedas läbivad fotoretseptori plasmamembraani. Na + ja K + kontsentratsioonigradiente hoitakse varda plasmamembraanil sisemise segmendi membraanis paikneva naatrium-kaaliumpumba aktiivse töö tõttu.
Välissegmendi membraanil tekkinud hüperpolarisatsiooniretseptori potentsiaal levib seejärel mööda rakku selle presünaptilisse otsa ja viib vahendaja (glutamaadi) vabanemiskiiruse vähenemiseni. Seega lõpeb fotoretseptori protsess neurotransmitteri vabanemise kiiruse vähenemisega fotoretseptori presünaptilisest otsast.
Mitte vähem keerukas ja täiuslik on mehhanism fotoretseptori algse pimeduse taastamiseks, st selle võimet reageerida järgmisele valgusärritusele. Selleks on vaja uuesti avada plasmamembraanis olevad ioonikanalid. Kanali avatud oleku tagab selle seos cGMP molekulidega, mis omakorda on otseselt tingitud vaba cGMP kontsentratsiooni suurenemisest tsütoplasmas. See kontsentratsiooni tõus tuleneb metarodopsiin II võime kadumisest transduktiiniga interaktsiooniks ja ensüümi guanülaattsüklaasi (GC) aktiveerimisest, mis on võimeline GTP-st cGMP-d sünteesima. Selle ensüümi aktiveerimine põhjustab kontsentratsiooni langust
vaba kaltsiumi tsütoplasmas membraani ioonikanali sulgumise ja valguvahetaja pideva töö tõttu, mis kaltsiumi rakust välja paiskab. Kõige selle tulemusena suureneb cGMP kontsentratsioon rakus ja cGMP seondub taas plasmamembraani ioonkanaliga, avades selle. Avatud kanali kaudu hakkavad Na + ja Ca 2+ uuesti rakku sisenema, depolariseerides retseptori membraani ja viies selle "pimedasse" olekusse. Depolariseeritud retseptori presünaptilisest otsast kiireneb taas vahendaja vabanemine.
võrkkesta neuronid. Võrkkesta fotoretseptorid on sünaptiliselt ühendatud bipolaarsete neuronitega (vt joonis 14.6, B). Valguse toimel väheneb vahendaja (glutamaadi) vabanemine fotoretseptorist, mis viib bipolaarse neuroni membraani hüperpolariseerumiseni. Sellest edastatakse närvisignaal ganglionrakkudesse, mille aksonid on nägemisnärvi kiud. Signaali ülekanne nii fotoretseptorist bipolaarsele neuronile kui ka sealt ganglionilisele neuronile puur toimub impulsiivsel viisil. Bipolaarne neuron ei tekita impulsse väga väikese vahemaa tõttu, mille kaudu ta signaali edastab.
Iga 130 miljoni fotoretseptori raku kohta on ainult 1 250 000 ganglionrakku, mille aksonid moodustavad nägemisnärvi. See tähendab, et paljude fotoretseptorite impulsid koonduvad (koonduvad) bipolaarsete neuronite kaudu ühte ganglionrakku. Ühe ganglionrakuga ühendatud fotoretseptorid moodustavad ganglionraku vastuvõtuvälja. Erinevate ganglionrakkude vastuvõtuväljad kattuvad osaliselt üksteisega. Seega võtab iga ganglionrakk kokku paljudes fotoretseptorites esineva ergastuse. See suurendab valgustundlikkust, kuid halvendab ruumilist eraldusvõimet. Ainult võrkkesta keskel, fovea piirkonnas, on iga koonus ühendatud ühe niinimetatud bipolaarse kääbusrakuga, millega on samuti ühendatud vaid üks ganglionrakk. See tagab siin kõrge ruumilise eraldusvõime, kuid vähendab järsult valgustundlikkust.
Võrkkesta naaberneuronite interaktsiooni tagavad horisontaalsed ja amakriinsed rakud, mille käigus levivad signaalid, mis muudavad sünaptilist ülekannet fotoretseptorite ja bipolaarsete rakkude (horisontaalsed rakud) ning bipolaarsete ja ganglionrakkude vahel (amakriinrakud). Amakriinrakud inhibeerivad külgnevaid ganglionrakke lateraalselt.
Lisaks aferentsetele kiududele on nägemisnärvis ka tsentrifugaalsed ehk eferentsed närvikiud, mis toovad ajust signaale võrkkestale. Arvatakse, et need impulsid toimivad võrkkesta bipolaarsete ja ganglionrakkude vahelistel sünapsidel, reguleerides nendevahelist ergastuse juhtivust.
Närvide rajad ja ühendused V visuaalne süsteem. Võrkkesta visuaalsest teabest piki nägemisnärvi kiude (II paar
kraniaalnärvid) tormab ajju. Iga silma nägemisnärvid kohtuvad aju põhjas, kus nad moodustavad osalise kiasma. Siin läheb osa iga nägemisnärvi kiududest tema enda silma vastasküljele. Kiudude osaline dekussioon annab igale ajupoolkerale informatsiooni mõlemast silmast. Need projektsioonid on korraldatud nii, et iga võrkkesta parempoolsete poolte signaalid sisenevad parema ajupoolkera kuklasagarasse ja võrkkesta vasakpoolsete poolte signaalid vasakusse poolkera.
Pärast optilist kiasmi nimetatakse nägemisnärve optilisteks traktideks. Need projitseeritakse mitmetesse ajustruktuuridesse, kuid põhiline arv kiude jõuab taalamuse subkortikaalsesse visuaalsesse keskusesse - lateraalsesse ehk välisesse geniculate body (NKT). Siit sisenevad signaalid visuaalse ajukoore esmasesse projektsioonialasse (vöötkoore ehk Brodmani järgi väljale 17). Kogu visuaalne ajukoor sisaldab mitut välja, millest igaüks täidab oma spetsiifilisi funktsioone, kuid võtab vastu signaale kogu võrkkestalt ja säilitab üldiselt oma topoloogia ehk retinotoopia (võrkkesta naaberpiirkondade signaalid sisenevad ajukoore naaberpiirkondadesse).
Visuaalse süsteemi keskuste elektriline aktiivsus.Electrichesky nähtused võrkkestas ja nägemisnärvis. Valguse toimel retseptorites ja seejärel võrkkesta neuronites tekivad elektrilised potentsiaalid, mis peegeldavad toimiva stiimuli parameetreid.
Võrkkesta kogu elektrilist reaktsiooni valgusele nimetatakse elektroretinogrammiks (ERG). Seda saab salvestada kogu silmast või otse võrkkestast. Selleks asetatakse üks elektrood sarvkesta pinnale ja teine - näonahale silma lähedal või kõrvapulgale. Elektroretinogrammil eristatakse mitmeid iseloomulikke laineid (joon. 14.8). Laine A peegeldab fotoretseptorite sisemiste segmentide (hilise retseptori potentsiaal) ja horisontaalsete rakkude ergastust. Laine b tekib võrkkesta gliaalrakkude (Mülleri) aktiveerimise tulemusena bipolaarsete ja amakriinsete neuronite ergastamisel vabanevate kaaliumiioonide poolt. Laine c peegeldab pigmendi epiteelirakkude aktiveerumist ja laine d - horisontaalsed rakud.
Valgusstiimuli intensiivsus, värvus, suurus ja kestus kajastuvad ERG-s hästi. Kõigi ERG lainete amplituud suureneb võrdeliselt valguse intensiivsuse logaritmiga ja ajaga, mille jooksul silm oli pimeduses. Laine d ( reaktsioon väljalülitamisele) on seda suurem, mida kauem tuli põles. Kuna ERG peegeldab peaaegu kõigi võrkkesta rakkude aktiivsust (v.a ganglionrakud), kasutatakse seda indikaatorit laialdaselt silmahaiguste kliinikus erinevate võrkkesta haiguste diagnoosimiseks ja ravi kontrollimiseks.
Võrkkesta ganglionrakkude ergastamine viib selleni, et mööda nende aksoneid (nägemisnärvi kiude) jõuab ajju
impulsid voolavad. Võrkkesta ganglionrakk on esimene "klassikalist" tüüpi neuron fotoretseptori-aju ahelas. Kirjeldatud on kolme peamist ganglionrakkude tüüpi: reageerivad sisselülitamisele (op-reaktsioon), väljalülitamisele (välja-reaktsioon) ja mõlemad (sisse-välja-reaktsioon) (joon. 14.9).
Võrkkesta keskosas olevate ganglionrakkude retseptsiooniväljade läbimõõt on palju väiksem kui perifeerias. Need vastuvõtlikud väljad on ümmargused ja kontsentriliselt ehitatud: ümmargune ergastav keskpunkt ja rõngakujuline pärssivat perifeerne tsoon või vastupidi. Retseptiivse välja keskel vilkuva valgustäpi suuruse suurenemisega suureneb ganglionraku reaktsioon (ruumiline summeerimine).
Lähedal asuvate ganglionrakkude samaaegne ergastamine põhjustab nende vastastikust inhibeerimist: iga raku vastused muutuvad väiksemaks kui ühe stimulatsiooni korral. See toime põhineb lateraalsel või lateraalsel inhibeerimisel. Naaberganglionrakkude vastuvõtlikud väljad kattuvad osaliselt, nii et samad retseptorid võivad osaleda vastuste genereerimises mitmelt neuronilt. Võrkkesta ganglionrakkude retseptiivsed väljad toodavad oma ümara kuju tõttu võrkkesta kujutise nn punkt-punkti kirjelduse: seda kuvab väga õhuke ergastatud neuronite mosaiik.
Elektrilised nähtused subkortikaalses nägemiskeskuses javisuaalne ajukoor. Ergastuspilt subkortikaalse nägemiskeskuse neuronaalsetes kihtides - välimine või lateraalne geniculate body (NKT), kuhu tulevad nägemisnärvi kiud, on paljudes aspektides sarnane võrkkestaga. Nende neuronite vastuvõtuväljad on samuti ümmargused, kuid väiksemad kui võrkkestas. Valgussähvatuse tagajärjel tekkivate neuronite reaktsioonid on siin lühemad kui võrkkesta puhul. Väliste genikulaarkehade tasandil toimub võrkkestast tulevate aferentsete signaalide interaktsioon nägemiskoorest tulevate eferentsete signaalidega, samuti kuulmis- ja muude sensoorsete süsteemide retikulaarse moodustumise kaudu. Need interaktsioonid tagavad sensoorse signaali kõige olulisemate komponentide valiku ja selektiivse visuaalse tähelepanu protsesside.
Välise genikulaarkeha neuronite impulsslahendused mööda nende aksoneid sisenevad ajupoolkerade kuklaosasse, kus asub visuaalse ajukoore esmane projektsiooniala (vöötkoore ehk väli 17). Siin on teabetöötlus palju spetsialiseeritum ja keerulisem kui võrkkesta ja väliste genikulaarkehade puhul. Nägemiskoore neuronitel ei ole ümmargused, vaid piklikud (horisontaalselt, vertikaalselt või ühes kaldus suunas) väikesed vastuvõtuväljad. Tänu sellele suudavad nad kogu pildist välja valida üksikuid ühe või teise orientatsiooni ja asukohaga joonte fragmente (orientatsioonidetektorid) ning reageerida neile valikuliselt.
Nägemiskoore igas väikeses piirkonnas, piki selle sügavust, on neuronid koondunud vaateväljas sama orientatsiooni ja vastuvõtuväljade lokaliseerimisega. Need moodustavad neuronite kolonni, mis kulgeb vertikaalselt läbi kõigi ajukoore kihtide. Veerg on näide sarnast funktsiooni täitvate kortikaalsete neuronite funktsionaalsest ühendusest. Nagu hiljutiste uuringute tulemused näitavad, võib nägemiskoores üksteisest kaugel olevate neuronite funktsionaalne ühinemine toimuda ka nende väljavoolude sünkroniseerimise tõttu. Paljud nägemiskoore neuronid reageerivad valikuliselt teatud liikumissuundadele (suunadetektorid) või mõnele värvile ning mõned neuronid reageerivad kõige paremini objekti suhtelisele kaugusele silmadest. Teavet visuaalsete objektide erinevate tunnuste (kuju, värv, liikumine) kohta töödeldakse paralleelselt ajukoore visuaalse tsooni erinevates osades.
Signaaliedastuse hindamiseks visuaalse süsteemi erinevatel tasanditel kasutatakse sageli kogu esilekutsutud potentsiaali (EP) registreerimist, mida loomadel saab samaaegselt eemaldada kõigist osakondadest ja inimestel - visuaalsest ajukoorest, kasutades peanahale kantud elektroode ( joon. 14.10).
Valgussähvatusest põhjustatud võrkkesta vastuse (ERG) ja ajukoore EP võrdlus võimaldab kindlaks teha patoloogilise protsessi lokaliseerimise inimese visuaalses süsteemis.
visuaalsed funktsioonid. valgustundlikkus. Absoluutne nägemistundlikkus. Visuaalse aistingu ilmnemiseks on vajalik, et valgusstiimulil oleks teatud minimaalne (lävi)energia. Valgusaistinguks vajalik minimaalne valguskvantide arv
et pimedas kohanemise tingimustes jääb vahemikku 8 kuni 47. Arvutatud on, et ühte varrast saab ergutada vaid 1 valguskvant. Seega on võrkkesta retseptorite tundlikkus valguse tajumiseks kõige soodsamates tingimustes füüsiliselt piirav. Võrkkesta üksikud vardad ja koonused erinevad veidi valgustundlikkuse poolest, kuid ühte ganglionrakku signaale saatvate fotoretseptorite arv on võrkkesta keskel ja perifeerias erinev. Koonuste arv võrkkesta keskel asuvas vastuvõtuväljas on umbes 100 korda väiksem kui võrkkesta perifeeria vastuvõtuväljas asuvate varraste arv. Vastavalt sellele on varraste süsteemi tundlikkus 100 korda suurem kui koonussüsteemil.
visuaalne kohanemine. Pimedusest valgusele üleminekul tekib ajutine pimedus ja seejärel väheneb järk-järgult silma tundlikkus. Seda visuaalse sensoorse süsteemi kohandamist ereda valguse tingimustega nimetatakse valgus kohanedamine. vastupidine nähtus (tume kohanemine) mida täheldatakse valgusküllasest ruumist peaaegu valgustamata ruumi liikumisel. Alguses ei näe inimene fotoretseptorite ja nägemisneuronite vähenenud erutatavuse tõttu peaaegu midagi. Järk-järgult hakkavad ilmnema objektide kontuurid ja siis erinevad ka nende detailid, kuna fotoretseptorite ja visuaalsete neuronite tundlikkus pimedas suureneb järk-järgult.
Valgustundlikkuse suurenemine pimedas viibimise ajal toimub ebaühtlaselt: esimese 10 minutiga suureneb see kümneid kordi ja seejärel tunni jooksul kümneid tuhandeid kordi. Olulist rolli selles protsessis mängib visuaalsete pigmentide taastamine. Koonuse pigmendid taastuvad pimedas kiiremini kui varras rodopsiin, seetõttu on pimedas viibimise esimestel minutitel kohanemine tingitud koonustes toimuvatest protsessidest. See esimene kohanemisperiood ei too kaasa suuri muutusi silma tundlikkuses, kuna koonusaparaadi absoluutne tundlikkus on madal.
Järgmine kohanemisperiood on tingitud rodopsiini taastamisest. See periood lõpeb alles esimese pimedas viibimise tunni lõpus. Rodopsiini taastamisega kaasneb varraste valgustundlikkuse järsk (100 000-200 000 korda) tõus. Tänu maksimaalsele tundlikkusele pimedas ainult varrastega, on nõrgalt valgustatud objekt nähtav ainult perifeerse nägemisega.
Kohanemises mängib olulist rolli lisaks visuaalsetele pigmentidele ka võrkkesta elementide vaheliste ühenduste muutus (lüliti). Pimedas suureneb ganglionraku vastuvõtuvälja erutuskeskuse pindala horisontaalse inhibeerimise nõrgenemise või eemaldamise tõttu. See suurendab bipolaarsete neuronite fotoretseptorite ja ganglionraku bipolaarsete neuronite lähenemist. Selle tulemusena suureneb võrkkesta perifeeria ruumilise summeerimise tõttu valgustundlikkus pimedas.
Kesknärvisüsteemi mõjust sõltub ka silma valgustundlikkus. Ajutüve retikulaarse moodustumise mõnede osade ärritus suurendab impulsside sagedust nägemisnärvi kiududes. Kesknärvisüsteemi mõju võrkkesta valgusega kohanemisele avaldub ka selles, et ühe silma valgustus vähendab valgustamata silma valgustundlikkust. Valgustundlikkust mõjutavad ka heli-, haistmis- ja maitsesignaalid.
Diferentsiaalne visuaalne tundlikkus. Kui valgustatud pinnal, mille heledus on I, rakendage lisavalgustust (dl), siis vastavalt seadusele




